| x | x | ||||
|
|
 |
||||
| BACTERIOLOGIE | IMMUNOLOGIE | MYCOLOGY | PARASITOLOGY | VIROLOGIE | |
|
|
|||||
|
|
|||||
|
Let us know what you think |
|||||
|
|
|||||
|
Logo image © Jeffrey Nelson, Rush University, Chicago, Illinois and The MicrobeLibrary |
|||||
|
READING: |
|||||
|
MOTS CLES |
Dans cette section, la structure des enveloppes
cellulaire des Gram négatives, Gram positives et acido-alcoolo-résistantes
seront discutées. La composition et la fonction de macromolécules uniques de
l’enveloppe cellulaire et leur biosynthèse sera décrite. De plus, les endospores,
qui sont inhabituels de diverses façons, (incluant la structure de la membrane
cellulaire) seront discutés. L’enveloppe cellulaire L’enveloppe cellulaire peut être définie comme constituée de la membrane de la cellule et de la paroi plus une membrane externe si elle est présente. La paroi consiste en une couche de peptidoglycane et de structures attachées. La plupart des enveloppes bactériennes sont réparties en deux catégories principales (Figure 1) : les Gram positives et les Gram négatives. D’autres types de parois sont retrouvés dans quelques espèces bactériennes (ni Gram positives ni Gram négatives). Le peptidoglycane (PG) est une macromolécule unique
en forme de sac, fortement réticulée qui entoure la membrane cellulaire
bactérienne et lui fournit une certaine rigidité. Il est très gros (des
milliards en terme de poids moléculaire; en comparaison de protéines qui ont des
poids moléculaires de l’ordre du millier). Le peptidoglycane consiste en un
squelette de glycane (polysaccharide) constitué d’acide N-acétyle muramique et
de N-acétyle glucosamine avec des chaines latérales de protéines contenant des
acides aminés sous forme D et L et dans certains cas de l’acide diaminopimélique.
Les chaînes latérales sont réticulées par des ponts peptidiques. La structure de
ces ponts peptidiques varie en fonction de l’espèce bactérienne. L’acide
muramique, l’acide D-amino et l’acide diaminopimélique ne sont pas synthétisés
par les mammifères. Le PG est retrouvé dans toutes les eubactéries excepté dans
Chlamydia et Mycoplasma.
|
||||
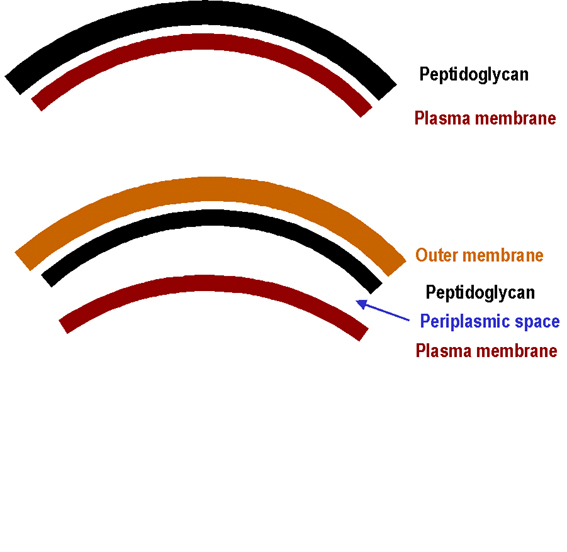 Figure 1 : Diagramme simplifié de l’enveloppe cellulaire de bactéries à Gram
positif et à Gram négatif.
Figure 1 : Diagramme simplifié de l’enveloppe cellulaire de bactéries à Gram
positif et à Gram négatif. |
Enveloppe cellulaire des Gram positifs
(Figure 2) L’acide téïchoique (dont le
squelette est généralement constitué de polymères de ribitol ou glycérol
contenant du phosphore) ou l’acide téïchurique (polysaccharides contenant de
l’acide glucuronique) sont liés de manière covalente au peptidoglycane. Ces
molécules chargées négativement seraient impliquées dans la concentration d’ions
métalliques à partir de l’environnement. Les acides téïchoiques peuvent aussi
diriger les enzymes autolytiques vers les sites de digestion du peptidoglycane (autolyse),
ceci étant une des étapes de la biosynthèse de la paroi. Dans certains cas des
polysaccharides neutres sont présents. L’acide lipotéïchoique, dans de
nombreuses bactéries, est généralement associé avec la membrane cellulaire. Dans
d’autres cas, il est retrouvé dans les fimbriae à l’extérieur de la cellule.
Quand il est exprimé à l’extérieur de la cellule il peut être impliqué dans
l’adhésion aux cellules épithéliales permettant ainsi la colonisation de la
gorge (ex : par les streptocoques du groupe A).
|
||||
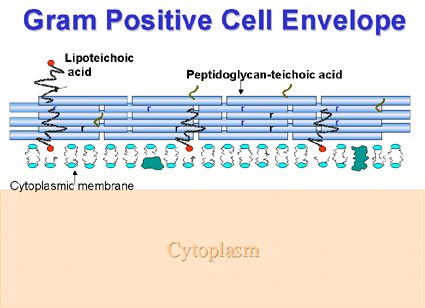 Figure
2 : Composant structuraux de l’enveloppe cellulaire d’un Gram positif. Figure
2 : Composant structuraux de l’enveloppe cellulaire d’un Gram positif.
|
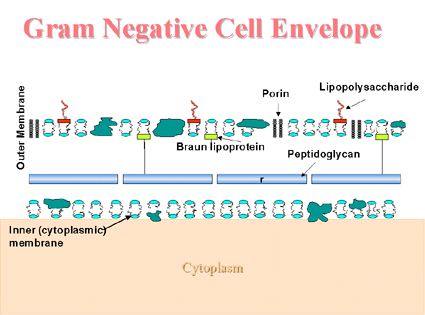
Enveloppe cellulaire des Gram négatifs
(Figure 3) Lié de manière covalente au fin peptidoglycane, on retrouve la lipoprotéine de Braun qui lie la membrane externe à l’enveloppe cellulaire. Comme les autres membranes, la membrane externe contient des protéines et des phospholipides. Contrairement aux autres membranes, elle contient des molécules supplémentaires, le lipopolysaccharide. Le lipopolysaccharide est important pour la cellule bactérienne car il procure une barrière perméable aux substances hydrophobes. Le lipopolysaccharide consiste en trois régions : l’antigène O externe, une partie intermédiaire et une région interne contenant le lipide A. La partie intermédiaire contient plusieurs sucres (heptoses et acide kétodeoxyoctonique), retrouvés nulle part ailleurs dans la nature, et le lipide A contient des bêtahydroxy acides gras (peu communs dans la nature). Cette molécule présente une activité d’endotoxine. Les porines présentes dans la membrane externe aident à la formation de canaux pour permettre le passage de petits nutriments hydrophiles (comme les sucres) à travers la membrane externe. Acid fast and related bacteria (mycobactéries, norcardia et corynebacteria) Les enveloppes cellulaires de ces organismes sont
beaucoup plus complexes que celles des autres bactéries. L’acide mycolique (chaînes
d’acides gras longues et ramifiées) est lié de manière covalente au
peptidoglycane via un polysaccharide. D’autres composés contenant de l’acide
mycolique et d’autres lipides complexes forment une couche membraneuse épaisse
et cireuse à l’extérieur de la couche de peptidoglycane.
|
||||
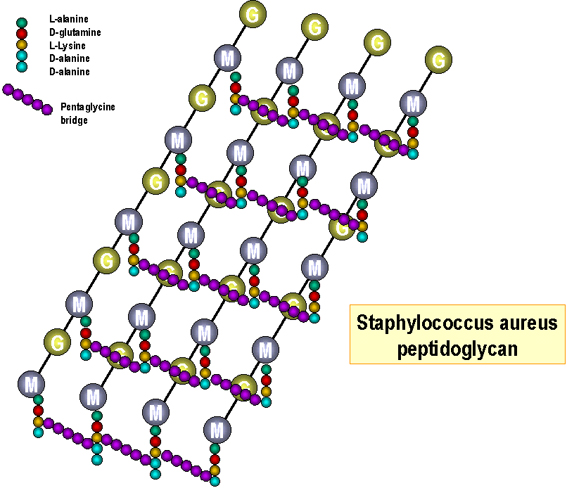
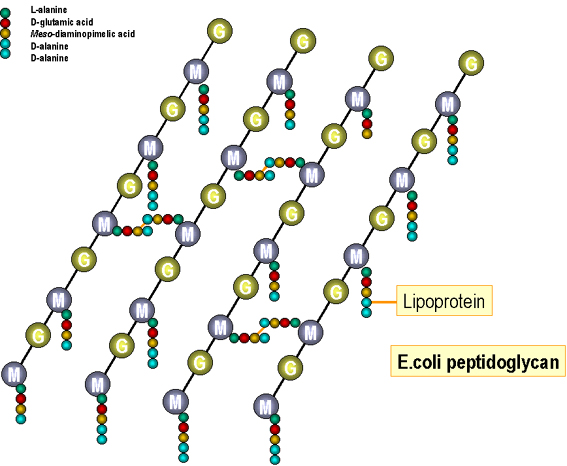 Figure 4 : Structure du peptidoglycane d’E. coli.
Figure 4 : Structure du peptidoglycane d’E. coli.
|
Synthèse de macromolécules de l’enveloppe
cellulaire Le peptidoglycane (Figures 4
et 5) : La sous-unité précurseur (le muramyl pentapeptide attaché à l’uridine
diphosphate, UDP) est synthétisée dans le cytoplasme et transférée à la membrane
cellulaire. La sous-unité est déplacée enzymatiquement du nucléotide au lipide
porteur (undécaprénole/bactoprénole) et transformé en sous-unité complète
(disaccharide pentapeptide attachés avec des ponts peptidiques). Les sous-unités
ainsi achevées sont ensuite exportées à la paroi. Après libération du monomère,
l’undécaprénole est remis en circulation dans la membrane cellulaire et
réutilisé. Le squelette de glycane de la paroi existante est dégradé
enzymatiquement (par les autolysines) pour permettre l’insertion des sous-unités
nouvellement synthétisées. Si ces enzymes sont trop réactives, la paroi est
dégradée et la forte pression osmotique de la cellule fait éclater la membrane
cytoplasmique tuant ainsi la cellule (« autolyse »). La réticulation des chaînes
latérales du peptide de la sous-unité insérée à la chaîne existante se produit
ensuite enzymatiquement (protéines liant la pénicilline). Les sous-unités
complétées d’acide téïchoïque et teïchurique sont également synthétisées dans la
membrane cellulaire (sur des lipide porteur) avant d’être transportés et insérés
dans la paroi existante.
|
||||
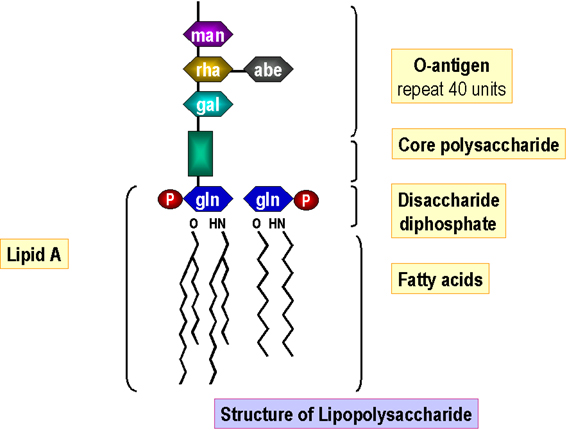 Figure 6 : Structure du lipopolysaccharide.
Figure 6 : Structure du lipopolysaccharide. |
Lipopolysaccharide
Le lipide A est assemblé dans la membrane cellulaire
et les sucres du noyau sont attachés séquentiellement. Les sous-unités de
l’antigène O sont synthétisées indépendamment (sur un lipide porteur comme dans
la synthèse du peptidoglycane). L’antigène O entièrement synthétisé est alors
attaché au noyau de lipide A (générant le lipopolysaccharide) dans la membrane
cellulaire avant le passage/ l’insertion dans la membrane externe (Figure 6).
|
||||
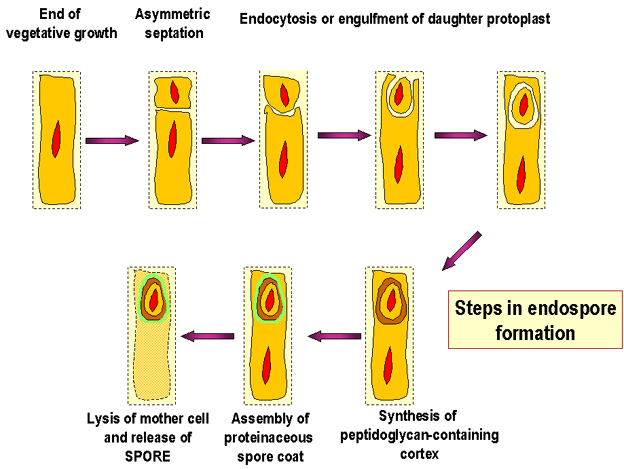 Figure 7 : Etapes de la formation d’une endospore.
Figure 7 : Etapes de la formation d’une endospore.
|
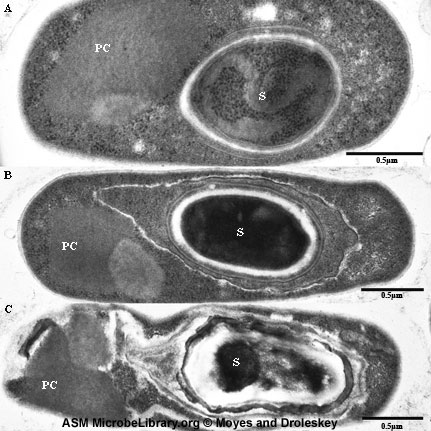
Endospores Ces cellules Gram positives modifiées ont une enveloppe cellulaire inhabituelle qui contient une membrane cellulaire et une membrane externe. La couche de peptidoglycane est moins réticulée que dans la plupart des cellules bactériennes et contient une forme déshydratée d’acide muramique. Le peptidoglycane de la spore est connu sous le nom de cortex et est retrouvé entre les deux membranes. Une épaisseur constituée de kératine fortement réticulée est retrouvée entourant l’extérieur de la cellule. La spore bactérienne est extrêmement résistante aux agents chimiques à cause de cette épaisseur. Normalement, au cours de la réplication bactérienne,
une cellule se divise, un septum se forme séparant la cellule mère en deux
cellules filles d’environ la même taille. Lorsque la sporulation se produit, la
division cellulaire est inégale et la « cellule mère » plus grosse enveloppe la
cellule fille. La membrane cellulaire de la cellule fille constitue la membrane
interne de la spore et la membrane cellulaire de la cellule mère forme la
membrane externe (Figure 7 et 8).
|
||||
|
|
|||||
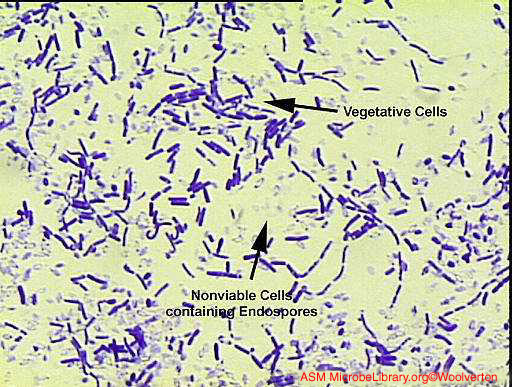 Figure 8A Bacillus showing vegetative and endospore-containing cells. Gram stain © Christopher J. Woolverton, Kent State University
Kent, Ohio and
The
MicrobeLibrary
Figure 8A Bacillus showing vegetative and endospore-containing cells. Gram stain © Christopher J. Woolverton, Kent State University
Kent, Ohio and
The
MicrobeLibrary
|
|||||
|
|
|
||||