| x | x | ||||
|
|
 |
||||
| BACTERIOLOGY | IMMUNOLOGY | MYCOLOGY | PARASITOLOGY | VIROLOGY | |
|
|
|
||||
|
|
|||||
|
Let us know what you think |
|||||
|
|
|||||
|
Logo image © Jeffrey Nelson, Rush University, Chicago, Illinois and The MicrobeLibrary |
|||||
|
|
|||||
|
KEY WORDS |
In this section, the structure of the Gram negative, Gram positive and acid fast cell envelopes will be discussed. The composition and function of unique cell envelope macromolecules and their biosynthesis will be described. In addition endospores, which are unusual in many ways (including cell envelope structure) will be discussed. Cell Envelope The cell envelope may be defined as the cell membrane and cell wall plus an outer membrane if one is present. The cell wall consists of the peptidoglycan layer and attached structures. Most bacterial cell envelopes fall into two major categories (Figure 1): Gram positive and Gram negative. This is based on Gram staining characteristics that reflect major structural differences between the two groups. Other types of cell wall are found in a few bacterial species (neither Gram positive nor Gram negative). The peptidoglycan is a single bag-shaped, highly cross-linked macromolecule that surrounds the bacterial cell membrane and provides rigidity. It is huge (billions in molecular weight; compare proteins which are thousands in molecular weight). Peptidoglycan consists of a glycan (polysaccharide) backbone consisting of N-acetyl muramic acid and N-acetyl glucosamine with peptide side chains containing D- and L- amino acids and in some instances diaminopimelic acid. The side chains are cross-linked by peptide bridges. These peptide bridges vary in structure among bacterial species. Muramic acid, D-amino acids and diaminopimelic acid are not synthesized by mammals. PG is found in all eubacteria except Chlamydia and Mycoplasma.
|
||||
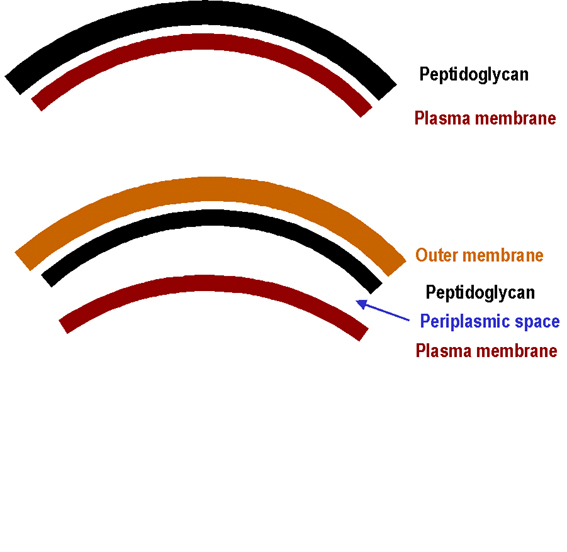 Figure 1. Simplified diagram of the cell envelope of Gram positive and Gram negative
bacteria
Figure 1. Simplified diagram of the cell envelope of Gram positive and Gram negative
bacteria |
Gram Positive Cell Envelope (Figure 2) Covalently bound to the thick peptidoglycan are teichoic acid (their backbones are usually phosphorus-containing polymers of ribitol or glycerol) or teichuronic acid (glucuronic acid- containing polysaccharides). These negatively charged molecules are believed to be involved in concentrating metal ions from the surroundings. Teichoic acids can also direct autolytic enzymes to sites of peptidoglycan digestion (autolysis), one of the steps in cell wall biosynthesis. In some instances neutral polysaccharides are present. Lipoteichoic acid, in many bacteria, is generally associated with the cell membrane. In other instances it occurs in the fimbriae on the outside of the cell. When expressed on the cell exterior it can be involved in adhesion to epithelial cells allowing colonization of the throat (e.g. by the group A streptococcus).
|
||||
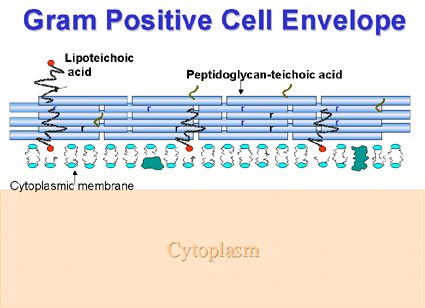 Figure
2. The component structures of the Gram positive cell envelope Figure
2. The component structures of the Gram positive cell envelope
|
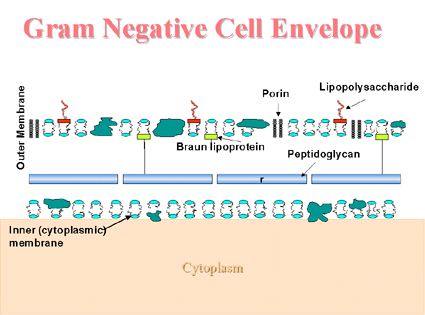
The Gram negative cell envelope (Figure 3) Covalently linked to the thin peptidoglycan is the Braun lipoprotein which binds the outer membrane to the cell wall. Like other membranes, the outer membrane contains proteins and phospholipids. Unlike other membranes, it contains additional molecules (lipopolysaccharide). The lipopolysaccharide is important to the bacterial cell since it helps to provide a permeability barrier to hydrophobic substances. The lipopolysaccharide consists of three regions: an outer O antigen, a middle core and an inner lipid A region. The core contains several sugars (heptoses and ketodeoxyoctonic acid), not found elsewhere in nature, and lipid A contains β hydroxyfatty acids (uncommon in nature). The molecule displays endotoxin activity. Porins in the outer membrane help form channels to allow passage of small hydrophilic nutrients (such as sugars) through the outer membrane.
The cell envelopes of these organisms are considerably more complex than other bacteria. Mycolic acid (long, branch chained fatty acids) covalently bound via a polysaccharide to peptidoglycan. Other mycolic acid-containing compounds and other complex lipids form a thick waxy membranous layer outside the peptidoglycan layer.
|
||||
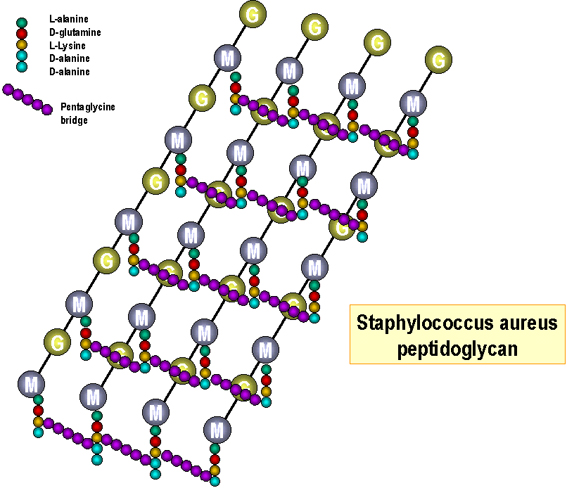
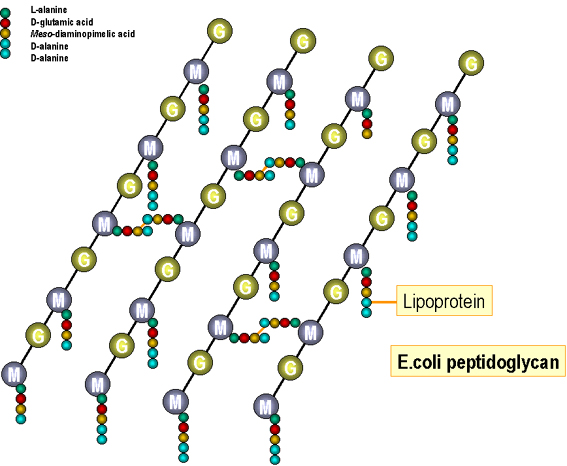 Figure 4. Structure of peptidoglycan of E. coli
Figure 4. Structure of peptidoglycan of E. coli
|
Synthesis of cell envelope macromolecules Peptidoglycan (Figures 4 and 5): The precursor subunit (muramyl pentapeptide attached to uridine diphosphate, UDP) is synthesized in the cytoplasm and passed to the cell membrane. The subunit is moved enzymatically from the nucleotide to a lipid carrier (undecaprenol/bactoprenol) and built into a completed subunit (disaccharide pentapeptide with attached bridge peptide). The completed subunits are then exported to the cell wall. After release of the monomer the undecaprenol is recirculated in the cell membrane and used again. The glycan backbones of the existing cell wall is enzymatically broken (by autolysins) to allow insertion of the newly synthesized subunit. If these enzymes are overactive, the cell wall becomes degraded and the high osmotic pressure of the cell bursts the cytoplasmic membrane killing the cell ("autolysis"). Cross-linking of the peptide side-chain of the inserted subunit to the existing chain then occurs enzymatically (penicillin binding proteins). Completed subunits of teichoic and teichuronic acids are also synthesized in the cell membrane (on lipid carriers) before transport and insertion into the existing cell wall.
|
||||
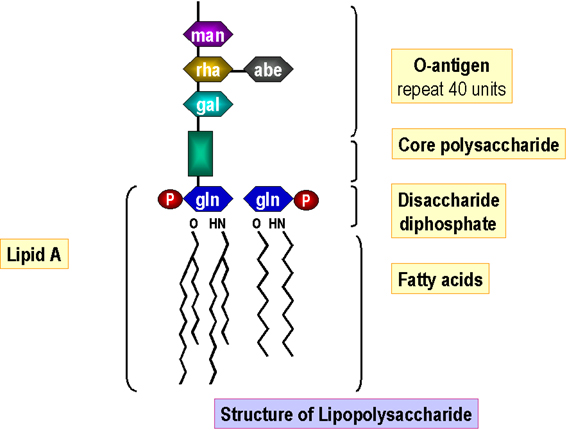 Figure 6. Structure of lipopolysaccharide
Figure 6. Structure of lipopolysaccharide |
Lipopolysaccharide Lipid A is assembled in the cell membrane and the core sugars attached sequentially. O-antigen subunits are independently synthesized (on a lipid carrier as in peptidoglycan synthesis). The fully synthesized O-antigen is then attached to the lipid A-core (generating lipopolysaccharide) in the cell membrane before passage/insertion into the outer membrane (Figure 6).
|
||||
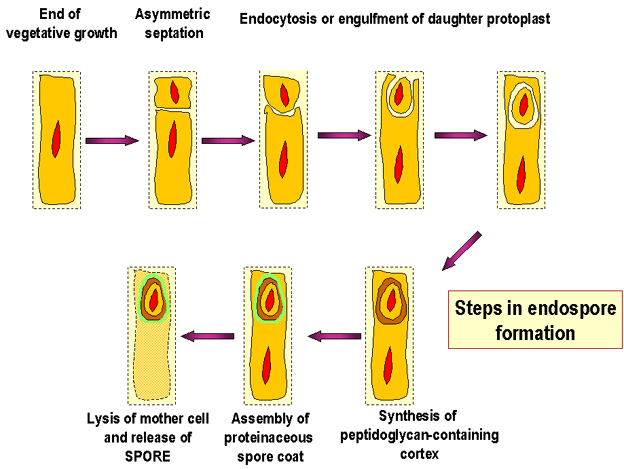 Figure 7. Steps in endospore formation
Figure 7. Steps in endospore formation
|
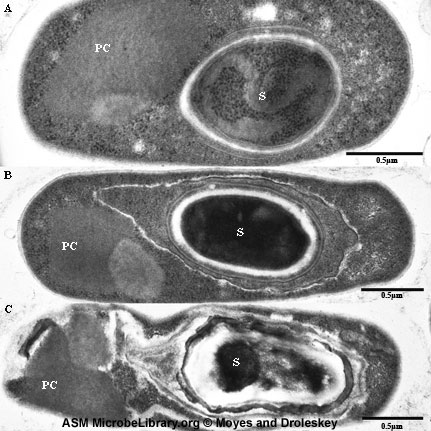
Endospores These modified Gram positive bacterial cells have an unusual cell envelope that contains a cell membrane and an outer membrane. The peptidoglycan layer is less cross-linked than in most bacterial cells and contains a dehydrated form of muramic acid. The spore peptidoglycan is referred to as a cortex and is found between the two membranes. A coat consisting of highly cross-linked keratin is found around the outside of the cell. The bacterial spore is highly resistant to chemical agents because of this coat. Normally in bacterial replication, as cells divide, a septum forms dividing the mother cell into two roughly equally sized daughters. When sporulation occurs, cell division is unequal and the larger so-called "mother cell" envelops the daughter cell. The cell membrane of the daughter cell constitutes the inner membrane of the spore and the cell membrane of the mother forms the outer membrane (Figure 7 and 8).
|
||||
|
|
|||||
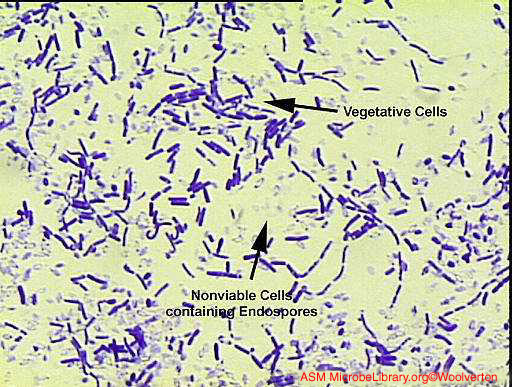 Figure 8A Bacillus showing vegetative and endospore-containing cells. Gram stain © Christopher J. Woolverton, Kent State University
Kent, Ohio and
The
MicrobeLibrary
Figure 8A Bacillus showing vegetative and endospore-containing cells. Gram stain © Christopher J. Woolverton, Kent State University
Kent, Ohio and
The
MicrobeLibrary
|
|||||
|
|
|
||||