| x | x | |||||||||||||||||||||||||||||||
|
|
 |
|||||||||||||||||||||||||||||||
| BACTERIOLOGIE | IMMUNOLOGIE | MYCOLOGY | PARASITOLOGY | VIROLOGIE | ||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||
| EN ANGLAIS | ||||||||||||||||||||||||||||||||
| IN SPANISH | ||||||||||||||||||||||||||||||||
| IN PORTUGUESE | ||||||||||||||||||||||||||||||||
| SHQIP - ALBANIAN | ||||||||||||||||||||||||||||||||
| IN FARSI | ||||||||||||||||||||||||||||||||
|
Let us know what you think |
||||||||||||||||||||||||||||||||
| SEARCH | ||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||
| Logo image © Jeffrey Nelson, Rush University, Chicago, Illinois and The MicrobeLibrary | ||||||||||||||||||||||||||||||||
|
Reading: Murray, 6th edition, Chapter 3 |
||||||||||||||||||||||||||||||||
|
MOTS CLES |
Les besoins des bactéries pour leur croissance incluent des sources d’énergie,
du carbone « organique » (ex : sucres et acides gras) et des ions métalliques
(ex : le fer). Une température optimale, le pH et les besoins (ou l’absence) en
oxygènes sont importants. Besoins en oxygène Les aérobies stricts doivent croître en présence d’oxygène ; ils ne peuvent pas réaliser la fermentation. Les anaérobies stricts de réalisent pas la phosphorylation oxydative. En outre, ils sont tués par l’oxygène ; il leur manque en effet certaines enzymes telles que la catalase [qui dégrade les ions peroxydes, H2O2, en eau et oxygène], la peroxydase [par laquelle NADH + H2O2 sont convertis en NAD et O2] et la superoxide dismutase [par laquelle le superoxide, O2., est converti en H2O2]. Ces enzymes détoxifient le peroxyde et les radicaux libres de l’oxygène produits durant le métabolisme en présence d’oxygène. Les anaérobies aérotolérantes sont des bactéries qui respirent de manière anaérobie, mais peuvent survivre en présence d’oxygène. Les anaérobies facultatives peuvent réaliser la fermentation et la respiration aérobie. En présence d’oxygène, la respiration anaérobie est généralement arrêtée et ces organismes respirent de manière aérobie. Les bactéries microaérophyles poussent bien en présence de faibles concentrations d’oxygène, mais sont tuées par de plus fortes concentrations. Besoins en nutriments Ceci inclut les sources de carbone organique, l’azote, le phosphore, le soufre et les ions métalliques comme le fer. Les bactéries secrètent de petites molécules qui lient le fer (des sidérophores comme l’entérobactine ou la mycobactine). Les sidérophores (liés au fer) sont ensuite internalisés par la cellule bactérienne via des récepteurs. L’hôte humain possède aussi des protéines de transport du fer (ex : la transferrine). Ainsi les bactéries qui rivalisent mal avec l’hôte pour le fer sont de faibles pathogènes. Température Les bactéries peuvent pousser dans des conditions de températures allant des points de congélation à quasiment l’ébullition de l’eau. Celles qui poussent le mieux entre les deux extrêmes sont appelées mésophiles ; ce qui inclus tous les pathogènes humains et bactéries opportunistes. Celles qui poussent de manière optimale aux plus basses et aux plus hautes des températures sont respectivement appelées psychrophiles et thermophiles. pH De nombreuses bactéries poussent mieux à un pH neutre ; cependant certaines
bactéries peuvent survivre et même pousser en conditions très acide ou alcaline.
|
|||||||||||||||||||||||||||||||
|
MOVIE Bacteria and Binary Fission |
||||||||||||||||||||||||||||||||
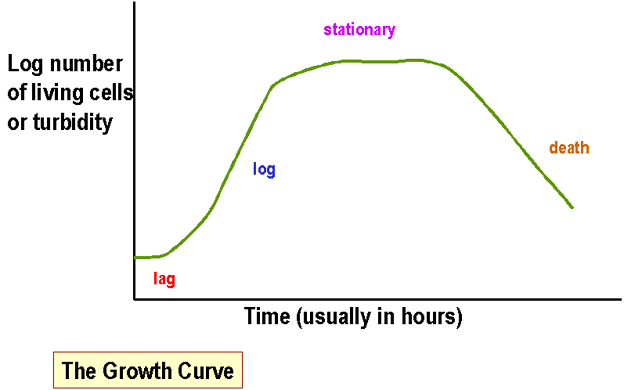 Figure 1 : Courbe de croissance
Figure 1 : Courbe de croissance |
Mesure de la masse bactérienne en culture
liquide de bactéries Les méthodes courantes incluent :
|
|||||||||||||||||||||||||||||||
|
|
METABOLISME DES SUCRES (comme exemple de voies métaboliques) Glycolyse (Voies de Embden, Meyerhof et Parnas [EMP]) C’est la voie la plus commune chez les bactéries pour le catabolisme des
sucres (également retrouvée dans la majorité des cellules animales et
végétales). Une série de processus enzymatiques résulte en la conversion de
sucres en pyruvate, générant de l’ATP (adénosine triphosphate) et du NADH (nicotinamide
adénine dinucléotide). L’énergie chimique nécessaire à ces fins
biosynthétiques est stockée dans le composé nouvellement formé (ATP ou NADH).
* Correspond au nombre de carbones dans la molécule
Respiration anaérobie La respiration anaérobie inclue la glycolyse et la fermentation. Durant
les dernières étapes de ce processus, le NADH (généré durant la glycolyse)
est converti en NAD en perdant un hydrogène. Cet hydrogène est ajouté au
pyruvate et, selon les espèces bactériennes, une variété de produits finaux
métaboliques est générée.
Respiration aérobie La respiration aérobie implique la glycolyse et le cycle de l’acide tricarboxylique (cycle de Krebs). Le pyruvate est entièrement dégradé en dioxyde de carbone (C1) au cours de ce processus, le NAD est converti en NADH. Ainsi, au cours de la fermentation aérobie, le NADH est généré à partir de deux sources (la glycolyse et le cycle de Krebs). La phosphorylation oxydative convertit l’excès de NADH en NAD et produit ainsi plus d’ATP (énergie stockée). Les ubiquinones et cytochromes sont des composants de la chaîne de transport des électrons impliquée dans le processus cité plus haut. La conversion de l’oxygène en eau est l’étape finale du processus.
|
|||||||||||||||||||||||||||||||
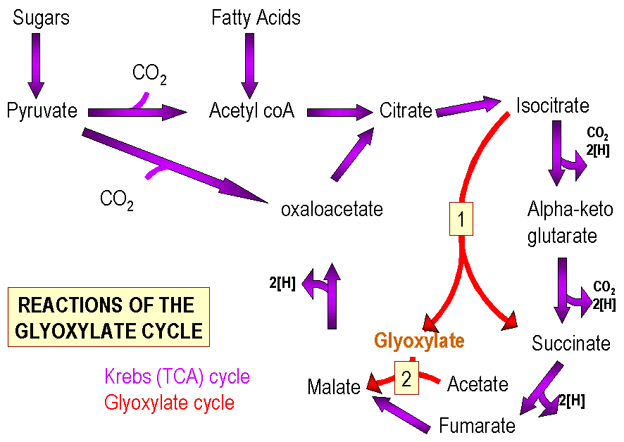 Figure 2 : Cycles de Krebs et du Glyoxylate
Figure 2 : Cycles de Krebs et du Glyoxylate |
Phosphorylation oxidative
Le cycle de Krebs (Figure 2) contient des intermédiaires à 4 ou 6 carbones. Le pyruvate (C3) peut alimenter le cycle de Krebs de telle manière que le nombre d’intermédiaires en C4/C6 reste le même ou augmente.
Ainsi, si certains composés du cycle sont retirés pour être utilisés dans d’autres voies biosynthétiques, ils peuvent être réapprovisionnés par cette réaction.
|
|||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||
|
|
METABOLISME DES ACIDES GRAS
Les acides gras sont dégradés en groupes acétyles (C2) qui approvisionnent le
cycle de Krebs par addition à un intermédiaire en C4 pour produire un C6.
Pendant le cycle, le C2 ajouté est perdu sous forme de CO2 et le C4 est régénéré.
Au total, il n’y a pas d’augmentation du nombre de molécules d’intermédiaires au
sein du cycle. Ainsi, si les acides gras sont la seule source de carbone, aucun
intermédiaire du cycle ne peut être retiré sans l’arrêter :
A la place, les bactéries utilisent le cycle du glyoxylate (Figure 2) (un cycle de Krebs modifié) au cours duquel les étapes enzymatiques, où deux molécules de CO2 sont retirées de l’intermédiaire en C6, sont esquivées. L’intermédiaire en C6 est converti en deux composés en C4 (tous deux des composants du cycle). Ainsi, pour chaque groupe acétyle (provenant d’acides gras), un intermédiaire supplémentaire du cycle est produit. La voie du glyoxalate n’est généralement pas trouvée dans les cellules animales puisque des acides gras préformés, présents dans la nourriture, sont utilisés.
En résumé, les fonctions du cycle de Krebs sont la biosynthèse et la production d’énergie. Cependant, si des intermédiaires sont retirés pour être utilisés dans d’autres voies métaboliques, ils doivent être remplacés. Le processus de remplacement pour l’utilisation des sucres et des acides gras est différent.
|
|||||||||||||||||||||||||||||||
|
|
|
|||||||||||||||||||||||||||||||