| x | x | ||||
|
|
 |
||||
| BACTERIOLOGY | IMMUNOLOGY | MYCOLOGY | PARASITOLOGY | VIROLOGY | |
|
|
|
||||
| En Español | |||||
| SHQIP - ALBANIAN | |||||
|
Let us know what you think FEEDBACK |
|||||
| SEARCH | |||||
|
|
|||||
|
THIS CHAPTER IS IN SEVERAL PARTS USE THE NEXT>> BUTTON ABOVE TO GO TO THE NEXT PART TO CONTINUE TO VIROLOGY CHAPTER EIGHT CLICK HERE |
|||||
|
LINKS TO OTHER HIV AND AIDS SECTIONS ARE AT THE BOTTOM OF THIS PAGE |
|||||
|
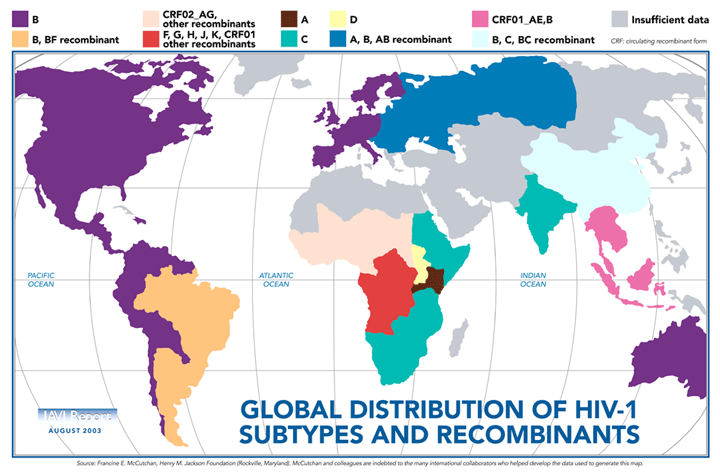
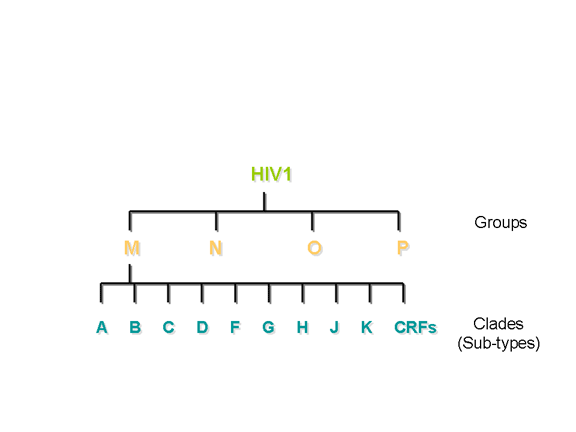
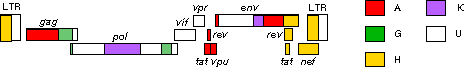
Two types of HIV can be distinguished genetically and antigenically. HIV-1 is the cause of the current worldwide pandemic while HIV-2 is found in west Africa but rarely elsewhere (figure 14a). HIV-2, which is transmitted in the same ways as HIV-1, causes AIDS much more slowly than HIV-1 but otherwise clinically the diseases are very similar. HIV-1 and HIV-2 are thought to have arisen from two natural hosts both harboring simian immunodeficiency virus (SIV): HIV-1 resulted from human infection by SIVcpz that infects chimpanzees and HIV-2 resulted from infection by SIVsmm harbored by sooty mangabeys. HIV-2 is closely related to the SIV found in west Africa. Sub-groups and sub-types There are three sub-groups of HIV-1, M (main or major), N (new) and O (outlier). Type O HIV-1 is mostly found in Cameroon and Gabon while the rare N sub-group is also found in Cameroon. It is very likely that SIVcpz infected humans on separate occasions to give rise to the three sub-groups. Based on nucleotide sequence analyses of the env and gag genes, it has been found that there are also at least ten different HIV-1 subtypes within the M group - these are designated A to J. The major one in North America, Latin America and the Caribbean, Europe, Japan and Australia is type B (figure 14a). Most sub-types are found in sub-Saharan Africa with A and D found at the highest rates in central and eastern Africa and C in southern Africa. Type C is also the predominant form in India and Nepal. It is type C that has caused the most infections worldwide. Type E is found in Thailand and central Africa, type F in Brazil and Romania, type G in Russia and Gabon, while type H is found in Zaire and in Cameroon. Subtype K is found in the Congo and Cameroon. Subtype I was a name given to an apparent sub-type found in Cyprus but the name is no longer used. In some countries, mosaics (recombinants) between different subtypes have been found (figure 14b). These arise when two different subtypes infect a person at the same time and recombination occurs. The former subtype I is a circulating recombinant form (or CRF) that is a recombinant of subtypes A, G, H and K (figure 14c and d). There is some evidence from laboratory studies that different HIV-1 subtypes can be transmitted by different routes. For example, type B found in western countries, may be transmitted more effectively by homosexual intercourse and via blood (as in intra-venous drug use) whereas types C and E may be transmitted more via a heterosexual route. This is because types C and E replicate better in Langerhans' cells found in the mucosa of the cervix, vagina and penis while type B replicates better in the rectal mucosa. It also appears that type E is more readily transmitted between sexual partners than type B. Subtype D seems to be more virulent than subtype A, that is infected persons progress to overt disease more rapidly. In addition subtypes D and C seem to be transmitted more effectively from mother to child than subtype A.
|
|||||
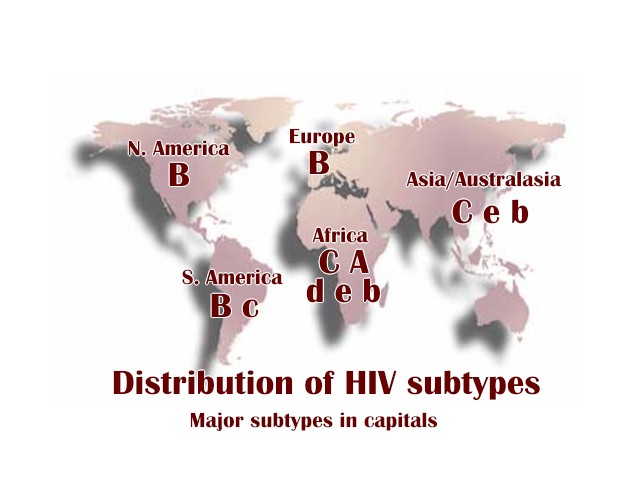 Figure 14a
Figure 14aHIV subtypes by continent
|
|||||
|
People who are infected but do not exhibit disease: Long-term non-progressors Ever since the AIDS epidemic began there have been people who are clearly exposed to the virus but who seem to exhibit no symptoms and normal CD4+ T cell counts. These are called long term non-progressors: People who have been infected with HIV for more than 7 years, who have stable CD4+ T cell counts above 600 per cu mm and have no history of symptoms and have not been taking anti-retroviral drugs. Their lymph node structure seems normal. The CD4+ T lymphocytes of these patients fall after primary infection and seroconversion but remain at normal levels thereafter, in some cases more than 15 years. This seems to be a heterogeneous group of people whose long-term non-progressive disease results from a robust CD8+ T cell immune response against HIV or a poorly replicative virus. About 5% of HIV-infected patients are long-term non-progressors.
People who are exposed but fail to become infected: Co-receptors and disease As noted below (see section 7 on co-receptors), a chemokine receptor on the surface of macrophages and activated T helper CD4+ cells was targeted by researchers as a co-receptor for HIV because of its known binding to three chemokines which seem to block infection. The nature of this co-receptor may be one explanation of the people who are exposed repeatedly to HIV but remain uninfected. It has been found that the cells of some exposed but uninfected individuals are very resistant to HIV infection because they have mutant chemokine receptors. The most common of these is a 32 base pair deletion that prevents surface expression of a chemokine receptor known as CCR5. CCR5 mutations are relatively rare. If two copies of the gene for CCR5 are defective, it is virtually impossible for virus to enter the cell and exposed patients are immune from HIV infection. Approximately 1 in 100 Caucasians have this double mutation. 17% have a single defective gene. No African-Americans have so far been found with a double mutation but about 2% have a single mutation. A single defective gene does not confer resistance but progress to disease is much less rapid. It is surprising that these people do not seem to have a reduced viral load or higher CD4 counts and so the reason why these people progress better is unclear. There are many of these chemokine receptors and some others are likely to be HIV co-receptors. A heterozygous mutation in CCR2 may defer AIDS by an average of 2 to 4 years. This protective CCR2 mutation is present in all races in the U.S. at a frequency of about 20 - 25%. One quarter of long term survivors are CCR2 or CCR5 mutants Other patients who do not become infected appear to make elevated levels of the chemokines that bind to these receptors and perhaps this keeps the receptor blocked. This might lead to therapy in which the chemokine receptors of macrophages and T-cells are blockaded - as noted below, some chemokines are powerful suppressors of HIV infection in vitro. An interesting fact is that one of the CCR5 mutations that confers almost complete resistance to infection by HIV (called “delta 32”) is thought to be the same mutation that rendered some people immune to the plague in the middle ages.
HLA antigens and IMMUNITY TO HIV disease There are other suggestions concerning individuals who are exposed to the virus without major apparent problems. A study of Nairobi prostitutes, repeatedly exposed to HIV (25% or more of their clients are HIV positive), has shown that many of these women have been free of disease for many years and seem to be completely resistant to infection. There seem to be associations between their resistance to infection and their class I and class II MHC (HLA) haplotypes. The strongest association of protection is with HLA-A*6802, A*0202 and B18. These women have mounted a very strong CTL response that is likely to mediate protection. It is possible that these particular class I MHC antigens allow a very efficient CTL response. Alternatively, they may present epitopes that are highly conserved between different HIV-1 variant strains. For example, one epitope to which there is a strong CTL response in these women is that presented by B18. This epitope is found to be located in a highly conserved part of HIV p24 protein. It appears to be conserved because it is very important in the assembly of the virus. Another important epitope is presented by HLA-A*6802 and this is in the protease. The protease may not be able to bear much mutation in this region without losing enzymic activity and so the virus cannot escape the immune response by mutation.
|
|||||
|
|
|||||
|
|||||
|
|
|||||