|
x |
x |
|
 |
 |
|
INFECTIOUS
DISEASE |
BACTERIOLOGY |
IMMUNOLOGY |
MYCOLOGY |
PARASITOLOGY |
VIROLOGY |
|
VIETNAMESE |
IMMUNOLOGY - CHAPTER SIX
GENETICS OF IMMUNOGLOBULINS
Gene Mayer, Ph.D
Emertius Professor of Pathology, Microbiology and Immunology
University of South Carolina
|
|
TURKISH |
|
FRANCAIS |
|
PORTUGUES |
|
SHQIP |
Let us know what you think
FEEDBACK |
|
SEARCH |
|
|

 |
|
Logo image
© Jeffrey Nelson, Rush University, Chicago, Illinois and
The MicrobeLibrary |
|
|
|
|
|
TEACHING
OBJECTIVES
To
describe the organization and expression of the immunoglobulin gene
families.
To
explain the origins of antibody diversity. |
History
Amino acid sequencing data
revealed that a single
C region could be associated with many different V
regions. Also, it was shown that a single
idiotype could be associated with
different C regions (eg. IgM and IgG). To explain these data it was
suggested that perhaps the two regions of the immunoglobulin molecule were coded for by
separate genes and that the V and C region genes were somehow joined before an
immunoglobulin molecule was made (i.e. there were two genes for one
polypeptide). This was a revolutionary concept but with the advent of
recombinant DNA technology, it has been shown to be the correct. The
immunoglobulin heavy
and light chains are coded for by three separate gene families each one on a
separate chromosome - one for the heavy chain and one for each of the light
chain types. Each of these gene families has several V region genes and one or
more C region genes. The V and C regions genes are not however immediately
adjacent to each other.
|
KEY WORDS
V
gene
C
gene
J
region
D
region
Leader
Enhancer
Promotor
Antibody
diversity
Germ line
theory
Somatic mutation theory
N region
insertions
Junctional diversity
Combinatorial association
Multispecificity
Clonal
selection |
Light
chain gene families
Germ line gene organization
The organization of the kappa and lambda light chain genes in the germ line
of undifferentiated cells is depicted in Figure 1.
-
Lambda light chains
The lambda gene family is composed of 4 C region genes, one for each subtype of
lamda chain, and approximately 30 V region genes. Each of the V region
genes is composed of two
exons, one (L) that codes for a leader region and
the other (V) that codes for most of the variable region. Upstream of each
of the C genes there is and additional exon called J (joining). The L, V, J
and C exons are separated by
introns (intervening non-coding sequences).
-
Kappa light chains
The kappa light chain gene family contains only one C region gene, since there
is only one type of kappa light chain. There are many V region genes
(approximately 250) each of which has a leader exon and a V exon. In the
κ gene family there are several J exons located between the V and C
genes. All of the exons are separated by introns.
|
 Figure 1
Figure 1
Organization of the kappa and lamda light chain genes in
the germ line or undifferentiated cells |
Gene rearrangement and
expression
As a cell differentiates into a mature B cell that will make
a light chain, there is a rearrangement of the various genes (exons) and the
gene begins to be expressed as depicted in Figure 2.
|
 Figure 2 Figure 2
As a cell differentiates into a mature B cell that will
make a light chain, there is rearrangement of the various genes (exons) |
As a cell commits to become a B
cell making a light chain, there is a rearrangement of the genes at the DNA
level such that one of the V genes is brought next to one of the J regions.
This occurs by a recombination event which removes the intron between the V
and J regions. The selection of which V gene is used is not totally random;
there is some preference for the use of V genes nearest to the J regions.
However, with time all V genes can be used so that all combinations of V genes
and J regions can be generated.
A consequence of this DNA
rearrangement is that the gene becomes transcriptionally active because a
promoter
(P), which is associated with the V gene, is brought close to an
enhancer (E), which is located in the intron between the J and C regions.
As transcription initiates from the promoter, a pre-mRNA is made which
contains sequences from the L, V J and C regions as well as sequences for
the introns between L and V and between J and C (Figure 2). This
pre-mRNA is processed (spliced) in the nucleus and the remaining introns
are removed. The resulting mRNA has the L, V J and C exons contiguous.
The mRNA is translated in the
cytoplasm and the leader is removed as the protein is transported into the
lumen of the endoplasmic reticulum. The light chain is assembled with a
heavy chain in the endoplasmic reticulum and the immunoglobulin is secreted via the
normal route of secretory proteins. The region V region of the mature
light chain is coded for by sequences in the V gene and J region and the C
region by sequences in the C gene.
|
 Figure
3
Figure
3
In addition to several J exons, the heavy chain gene
family also contains several additional exons called the D (diversity)
exons. All of the exons are separated by introns |
Heavy
chain gene family
Germ line gene
organization
The organization of the heavy chain genes is depicted
in Figure 3.
In the heavy chain gene
family there are many C genes, one for each class and subclass of
immunoglobulin. Each
of the C genes is actually composed of several exons, one for each domain
and another for the hinge region. In the heavy chain gene family there are
many V region genes, each composed of a leader and V exon. In addition to
several J exons, the heavy chain gene family also contains several
additional exons called the D (diversity) exons. All of the exons are
separated by introns as depicted in Figure 3.
|
| |
Gene rearrangements and
expression
As a cell differentiates into a mature B cell that will
make a heavy chain, there is a rearrangement of the various genes segments
(exons) and the gene begins to be expressed as depicted in Figures 4 and
5.
|
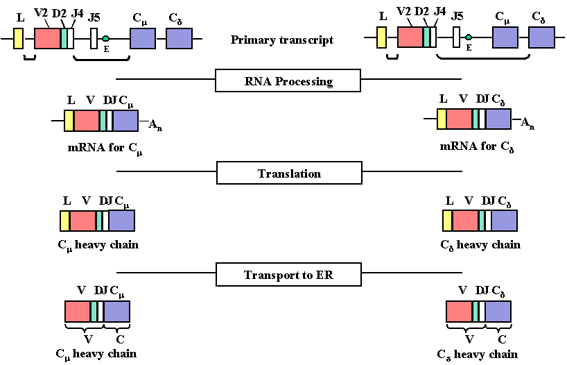
 Figure 4
Figure 4
As transcription initiates from the promoter a pre-mRNA is
made which contains sequences from the L, V, D, J Cμ and Cδ regions as
well as sequences for the introns between L and V, between J and Cμ, and
between Cμ and Cδ
 Figure
5
Figure
5
The pre-mRNA is processed (spliced) in the nucleus and the
remaining introns, including those between the exons in the C genes, are
removed |
As a cell commits to become a
B cell making a heavy chain, there are two rearrangements at the DNA
level. First, one of the D regions is brought next to one of the J regions
and then one of the V genes is brought next to the rearranged DJ region.
This occurs by two recombination events which remove the introns between
the V, D and J regions. As with the light chains the selection of the
heavy chain V gene is not totally random but eventually all of the V genes
can be used.
A consequence of these DNA
rearrangements is that the gene becomes transcriptionally active because a
promoter (P), which is associated with the V gene, is brought close to an
enhancer (E), which is located in the intron between the J and Cmu
regions. As transcription initiates from the promoter a pre-mRNA is made
which contains sequences from the L, V, D, J Cmu and Cdelta
regions as well as sequences for the introns between L and V, between J
and Cmu, and between Cmu and Cdelta
(Figure 4).
The pre-mRNA is processed
(spliced) in the nucleus and the remaining introns, including those
between the exons in the C genes, are removed (See Figure 5). The pre-mRNA
can be processed in two ways, one to bring the VDJ next to the Cmu
gene and the other to bring the VDJ next to the Cdelta gene.
The resulting mRNAs have the L, V, D, J and Cmu or Cdelta
exons contiguous and will code for a mu and a delta chain,
respectively.
The mRNAs are translated in
the cytoplasm and the leader is removed as the protein is transported into
the lumen of the endoplasmic reticulum. The heavy chain is assembled with
a light chain in the endoplasmic reticulum and the immunoglobulin is secreted via the
normal route of secretory proteins. The V region of the mature
heavy chain is coded for by sequences in the V gene, D region and J region
and the C region by sequences in the C gene.
|
 Figure
6
Figure
6
Flanking the V, J and D exons there are unique sequences
referred to as recombination signal sequences (RSS), which function in
recombination. Each RSS consists of a conserved nonamer and a conserved
heptamer that are separated by either 12 or 23 base pairs |
Mechanism
of DNA rearrangements
Flanking the V, J and D exons,
there are unique sequences referred to as recombination signal sequences (RSS),
which function in recombination. Each RSS consists of a conserved nonamer and
a conserved heptamer that are separated by either 12 or 23 base pairs (bp) as
illustrated in Figure 6. The 12bp and 23 bp spaces correspond to one or two
turns of the DNA helix.
Recombination only occurs between
a 1 turn and a 2 turn signal. In the case of the λ light chains there is
a 1 turn signal upstream of the J exon and a 2 turn signal downstream of Vlambda.
In the case of the κ light chains there is a 1 turn signal downstream of
the Vkappa gene and a 2 turn signal upstream of the J exon. In
the case of the heavy chains there are 1 turn signals on each side of the D exon and a 2 turn signal downstream of the V gene and a 2 turn signal upstream
of the J exon. Thus, this ensures that the correct recombination events will
occur.
The recombination event results
in the removal of the introns between V and J in the case of the light chains
or between the V, D, and J in the case of the heavy chains. The recombination
event is catalyzed by two proteins, Rag-1 and Rag-2. Mutations in the genes
for these proteins results in a
severe combined immunodeficiency disease (both
T and B cells are deficient), since these proteins and the RSS are involved in
generating both the B and T cell receptors for antigen.
|
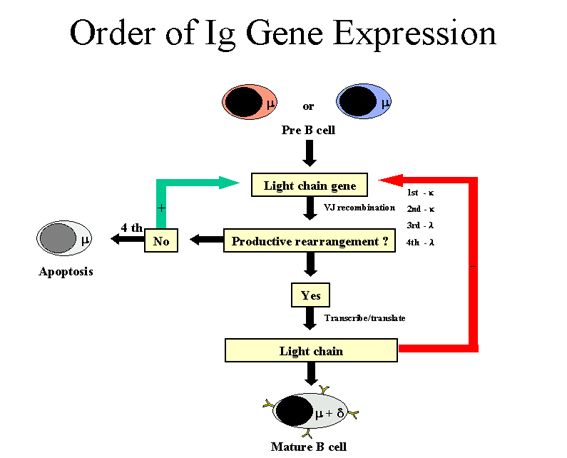
 Figure 7
Order of Ig gene
expression - Heavy chain
Figure 7
Order of Ig gene
expression - Heavy chain
 Figure 8
Figure 8
Order of Ig gene
expression - Light chain |
Order
of gene expression in Immunoglobulin gene families
An individual B cell only
produces one type of light chain and one class of heavy chain. (N.B.
The one exception is that a mature B cell can produce both μ and δ
heavy chains but the antibody specificity is the same since the same VDJ
region is found on the μ and δ chains). Since any B cell has both
maternal and paternal chromosomes which code for the immunoglobulin genes there must be
some orderly way in which a cell expresses its immunoglobulin genes so as to ensure that
only one type of light chain and one class of heavy chain is produced.
The order in which the immunoglobulin genes are
expressed in a B cell is depicted in Figure 7 and 8.
Heavy chain
(Figure 7)
A cell first attempts to rearrange one of its heavy chain genes;
in some cells the maternal chromosome is selected and in others the paternal
chromosome is selected. If the rearrangement is successful so that a heavy
chain is made, then no further rearrangements occur in the heavy chain genes.
If, on the other hand, the first attempt to rearrange the heavy chain genes is
unsuccessful (i.e. no heavy chain is made), then the cell attempts to
rearrange the heavy chain genes on its other chromosome. If the cell is
unsuccessful in rearranging the heavy chain genes the second time, it is
destined to be eliminated.
|
| |
Kappa light chain
(Figure 8)
When a cell successfully rearranges a heavy chain gene, it then
begins to rearrange one of its kappa light chain genes. It is a random event
whether the maternal or paternal kappa light chain genes are selected. If the
rearrangement is unsuccessful (i.e. it does not produce a functional
kappa light chain), then it attempts to rearrange the kappa genes on the
other chromosome. If a cell successfully rearranges a kappa light chain gene,
it will be a B cell that makes an immunoglobulin with a kappa light chain.
|
| |
Lambda light chain
(Figure 8)
If a cell is unsuccessful in rearranging both of its kappa light
chain genes, it then attempts to make a lambda light chain. It is a random
event whether the maternal or paternal lambda light chain genes are selected.
If the rearrangement is unsuccessful (i.e. it does not produce a
functional lambda light chain), then it attempts to rearrange the
lambda genes
on the other chromosome. If a cell successfully rearranges a lambda light
chain gene, it will be a B cell that makes an immunoglobulin with a lambda light chain.
The orderly sequence of
rearrangements in the immunoglobulin gene families explains:
-
Why an individual B cell
can only produce one kind of immunoglobulin with one kind of heavy and one
kind of light chain.
-
Why a individual B cell
can only make antibodies of one specificity.
-
Why there is allelic
exclusion in immunoglobulin allotypes at the level of an individual
immunoglobulin molecule but
co-dominant expression of allotypes in the organism as a whole.
|
 Figure 9
Figure 9
Origin of antibody diversity current concepts |
Origin of antibody diversity
Background
Antibody
diversity refers to the sum total of all the possible antibody specificities that
an organism can make. It is estimated that we can make 107 - 108
different antibody molecules. One of the major questions in immunology has been
how can we make so many different antibody molecules. Theories which have
attempted to explain the origin of antibody diversity fall into two major
categories.
Germ line theory
This theory states that we have a different V region gene for each
possible antibody we can make.
Somatic mutation theory
This theory states that we have only one or a few V region genes and the
diversity is generated by somatic mutations which occur in these genes.
Current Concepts
Our current thinking is that both the germ line and somatic mutation
theories have some merit. It is thought that antibody diversity is generated
by the following mechanisms.
1. A large
number of V genes
There are:
a) 30 lambda V genes
b) 300 kappa V genes
c) 1000 heavy chain V
genes
2. V-J and V-D-J joining
The region where the light chain V gene and J region or the heavy chain
V gene and D and J regions come together is in the third hypervariable
region. Since it is random which V and which J or D regions come together,
there is a lot of diversity that can be generated by V-J and V-D-J
joining.
3. Junctional diversity
(Inaccuracies in V-J and V-D and D-J recombination) - (Figure 9)
|
| |
Recombination between V-J and
V-D-J is not always perfect and additional diversity can arise by errors
that occur in the recombination event that brings the V region next to the
J or D regions or the D region next to the J region. It is estimated that
these inaccuracies can triple the diversity generated by V-J and V-D-J
joining. The diversity generated by this mechanism is occurring in the
third hypervariable region and thus, is directly affecting the combining
site of the antibody.
4. N region insertion
At the junction between D and J segments there is often an insertion of
a series of nucleotides which is catalyzed by the enzyme terminal
transferase. Terminal transferase catalyzes the random polymerization of
nucleotides into DNA without the need for a template. This leads to
further diversity in the third hypervariable region.
5. Somatic Mutation
There is evidence that somatic mutations are occurring in the V gene,
particularly in the place that codes for the second hypervariable region.
Thus, somatic mutation probably contributes to antibody diversity to some
extent.
6. Combinatorial
Association
Any individual B cell has the potential to make any one
of the possible heavy chains and any one of the possible light chains.
Thus, different combinations of heavy and light chains within an
individual B cell adds further diversity.
7. Multispecificity
Due to cross reactions between antigenic determinants of similar structure
an antibody can often react with more than one antigenic determinant. This
is termed multispecificity. Multispecificity also contributes to antibody
diversity.
An example of how these
mechanisms can generate a great deal of diversity is illustrated below:
|
| |
|
|
B Cell
Receptor (Immunoglobulin) |
|
Heavy |
Kappa |
|
V gene segments |
1000 |
300 |
|
D gene segments |
15 |
- |
|
J gene segments |
4 |
4 |
|
N region insertion |
++ |
- |
|
Junctional diversity |
+++ |
+ |
|
Somatic mutation |
+ |
+ |
|
Combinatorial association |
V x D x J
1000 X 15 X 4 |
V x J
300 x 4 |
|
Total |
6 x 104 |
1.2 x 103 |
|
|
|
|
Combinatorial association |
7.2 x 107 |
|
| |
These calculations do not take
into consideration the contributions of lambda light chains, somatic
mutation junctional diversity, N region insertions or multispecificity.
The process of gene
rearrangement of the heavy and light chains and the combinatorial
association of these chains occurs during B cell development and is independent
of antigen. Clones of B cells expressing all of the possible
antibody specificities are produced during development and antigen simply
selects those clones which have the appropriate receptor. The selected
clones are then activated, proliferate and differentiate into antibody
secreting plasma cells.
T Cell receptor for
antigen
T cells also have a receptor for
antigen on their surfaces. This receptor is not an immunoglobulin molecule but
it is composed of two different polypeptide chains which have constant and
variable regions analogous to the immunoglobulins. Diversity in the T cell
receptor is also generated in the same way as described for antibody diversity
(e.g. by VJ and VDJ joining of gene segments and combinatorial
association). However, no somatic mutation has been observed in T cells.
|
|
|
 Return to the Immunology Section of Microbiology and Immunology On-line
Return to the Immunology Section of Microbiology and Immunology On-line
This page last changed on
Wednesday, August 30, 2017
Page maintained by
Richard Hunt
|
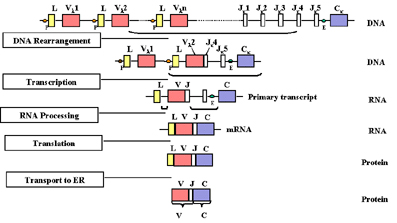 Figure 2
Figure 2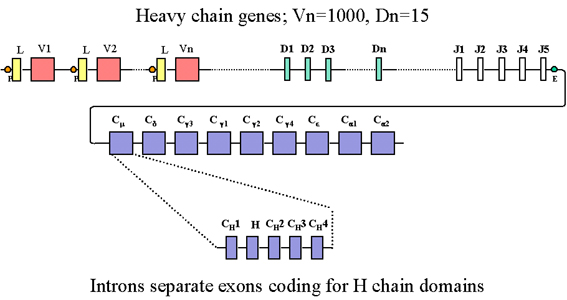 Figure
3
Figure
3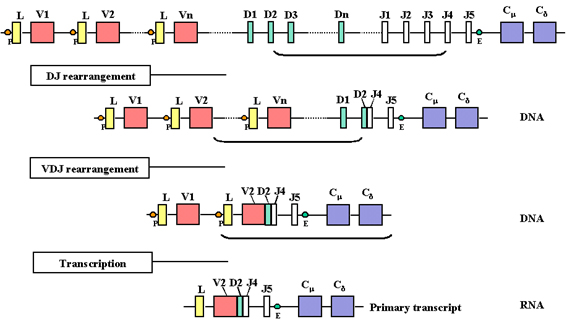 Figure 4
Figure 4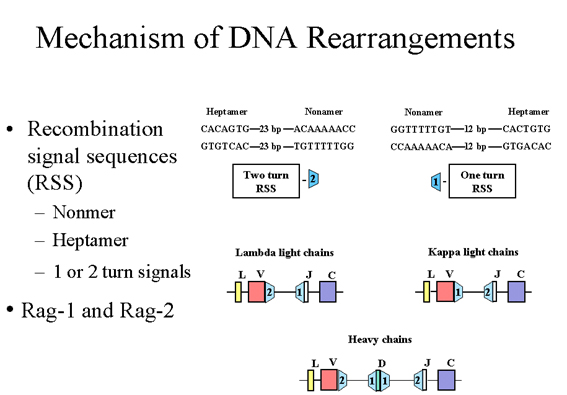 Figure
6
Figure
6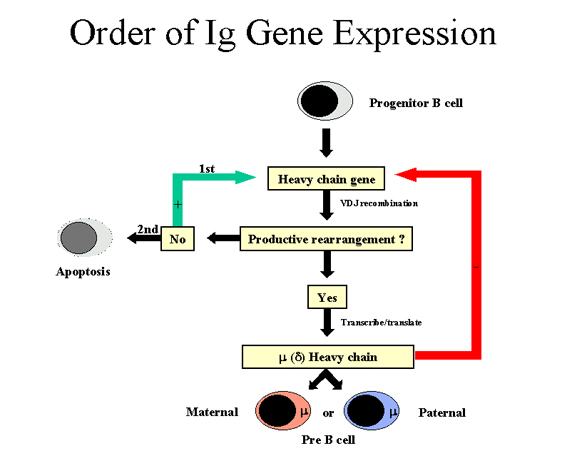 Figure 7
Order of Ig gene
expression - Heavy chain
Figure 7
Order of Ig gene
expression - Heavy chain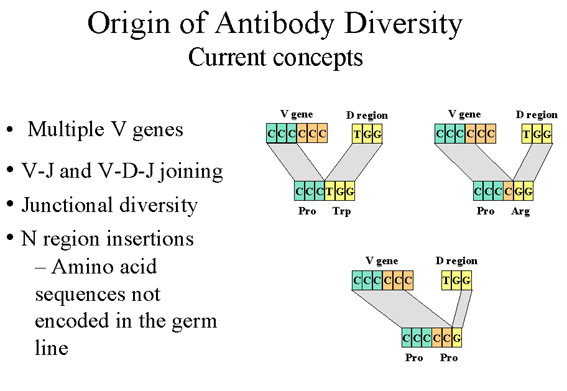 Figure 9
Figure 9