![]()

BACTERIOLOGIE – CHAPITRE UN
LA CELLULE BACTERIENNE
Alvin Fox, PhD
University of South Carolina School of Medicine
Columbia SC
USA
Emilie Camberlein, PhD
Maître de conférence en Biochimie
Universite de Nantes
Faculte des Sciences et des Techniques
Let us know what you think
FEEDBACK
![]()
Logo image © Jeffrey Nelson, Rush University, Chicago, Illinois and The MicrobeLibrary
LECTURES PROPOSEES
Murray et al., Microbiology
6th Ed., Chapter 2
MOTS CLES
Procaryote
Eubactéries (Bactéries)
Archéobactéries (Archaea)
Eucaryotes
Plasmide
Chromosome
Ribosome
Peptidoglycane (muréine, mucopeptide)
Coloration de Gram
Gram positif
Gram négatif
Enveloppe cellulaire
Membrane cellulaire
Paroi
Membrane externe
Espace périplasmique
Phosphorylation oxydative
Sphèroplaste/protoplaste
Flagelle
Chimiotaxie
Filament axial
Protéines périplasmiques de liaison
Perméases
Granules de stockage
Pilus (fimbriae)
Capsule (couche de slime, glycocalyx)
Endospore
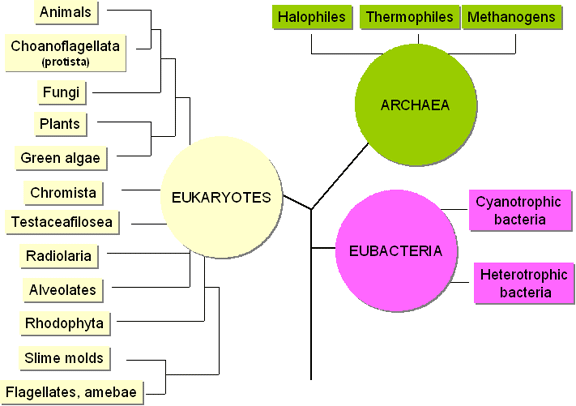 Figure 1a Les domaines du vivant.
Figure 1a Les domaines du vivant.
PROCARYOTES ET EUCARYOTES
Les « vraies » bactéries (ce qui inclut toutes les bactéries infectant l’homme) sont des membres d’un seul règne (les eubactéries, bactéries). Par ailleurs, un groupe d’organismes souvent retrouvés dans des environnements extrêmes forment un second règne (archéobactéries, Archaea). Morphologiquement, les deux règnes d’organismes semblent similaires, particulièrement du fait de l’absence de noyau, et sont donc classés ensemble en tant que procaryotes. Cependant, ils présentent des différences biochimiques majeures. La plupart des archaeas vit dans des environnements tels que les sources chaudes soufrées où elles sont exposées à des températures aussi hautes que 80°C et un pH de 2. Elles sont alors appelées thermoacidophiles. D’autres vivent dans des environnements contenant du méthane (méthanogènes) ou de fortes concentrations en sel (halophiles extrêmes).
Archaea
En se basant que les similarités de séquence d’ADN, il apparait que les branches archaea et eucaryote divergent des eubactéries avant de diverger l’une de l’autre (figure 1a) et d’une certaine manière, les archaeas sont biochimiquement plus proches des eucaryotes que des eubactéries. Par exemple, l’ARN polymérase des archaeas est aussi complexe, en termes de nombre de sous-unités, que les polymérases nucléaires des eucaryotes et il existe une homologie considérable en acides aminés avec certaines sous-unités eucaryotes. La structure des promoteurs des gènes chez les archaeas est également plus proche de celle des eucaryotes que des eubactéries mais, comme les eubactéries, les archaeas possèdent des opérons et les transcrivent en ARNm polycistroniques. Des similarités existent également entre les facteurs de synthèse des protéines des archaeas et des eucaryotes, suggérant que les mécanismes globaux de synthèse des protéines des eucaryotes et des archaeas pourraient être similaires. Les ARNs 16S des eubactéries et des archaeas présentent des séquences relativement distinctes.Les eubactéries (avec pour exception les genres Mycoplasma et Chlamydia) produisent du peptidoglycane (synonymes : muréine, mucopeptide, squelette de la paroi cellulaire). Le peptidoglycane contient un sucre unique, l’acide muramique, retrouvé nulle part ailleurs dans la nature. Les archéobactéries possèdent une pseudo-muréine dont la structure est différente de celle de la muréine des eubactéries.
Au vue du nombre croissant de similarités entre les archaeas et les eucaryotes, le terme archéobactéries n’est plus utilisé. Pour toutes les formes cellulaires de vie (dont les plantes, les animaux et les champignons) on fait communément référence aux eucaryotes.Les membres des Archaea ne sont pas des pathogènes humains et ne seront pas approfondis.
|
Similarités entre les Archaea et les Eucaryotes |
|||
| Eubacteria | Archaea | Eukaryotes | |
| Noyau | Non | Non | Oui : lié à la membrane |
| Nucléosomes/histones | Non | Oui | Oui |
| Opérons, ARNm polycistronique | Oui | Oui | Non |
| Introns | Non | Non | Oui |
| Protéines liant la boîte TATA | Non | Oui | Oui |
| Organites | Non | Non | Oui : mitochondrie, lysosomes, réticulum endoplasmique etc… |
| Chromosomes | Un circulaire | Un circulaire | Plus de un |
| ARN polymérase | Un (simple) | Plus de un (complexe) | Plus de un (complexe) |
| Acide aminé initiateur de protéine | N-formyl-méthionine | Méthionine | Méthionine |
| Sensibilité de la synthèse protéique à la toxine diphtérique | Insensible | Sensible | Sensible |
| Peptidoglycan | Oui | Non | Non |
| Synthèse protéique |
des Archaea sont plus proches de celles des eucaryotes que eubacteria |
||
DIFFERENCES PROCARYOTES/EUCARYOTES
La cellule procaryote, contrairement à la cellule eucaryote, n’est pas
compartimentée. Les membranes nucléaires, mitochondriales, du réticulum
endoplasmique, du Golgi, des phagosomes et lysosomes ne sont pas présentes
(Figures 1b, 2 et 3). Les procaryotes possèdent généralement un seul chromosome
circulaire. Puisqu’ils n’ont pas de membrane nucléaire, le chromosome est lié à
un site spécifique de la membrane cellulaire – le mésosome. Les ribosomes
procaryotes sont des 70S (S signifie unité Svedberg, une mesure de la taille),
alors que les ribosomes eucaryotes sont plus gros (80S). Les sous-unités
ribosomales procaryotes sont le 30S et le 50S (celles des eucaryotes sont plus
grosses). Le ribosome 30S contient l’ARN 16S, alors que le ribosome 50S contient
les ARN 23S et 5S. L’ARN ribosomal est plus large chez les eucaryotes (ex : ARNr
18S versus 16S). Les membranes bactériennes ne contiennent généralement pas de
stérols (ex : cholestérol).
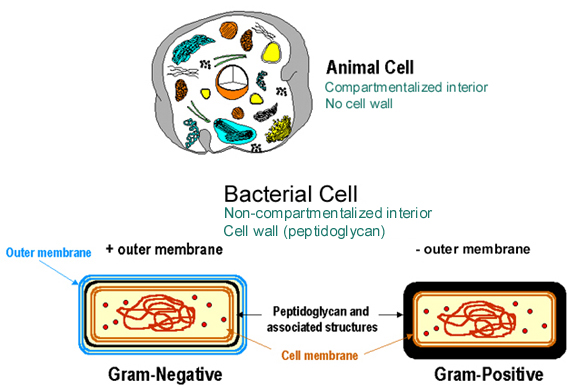 Figure 1b
Figure 1b Comparaison des eucaryotes et des procaryotes « eubactériens ».
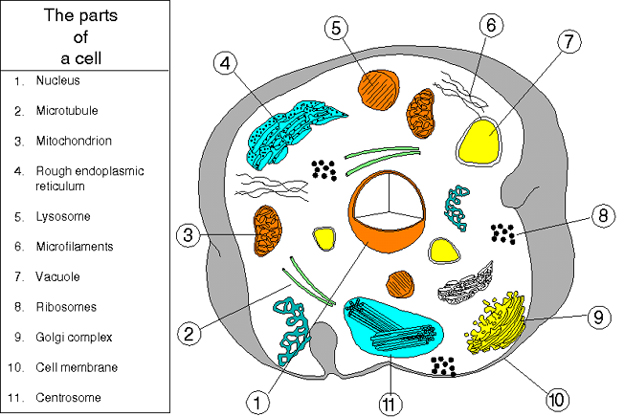 Figure
2
Figure
2
Une cellule animale
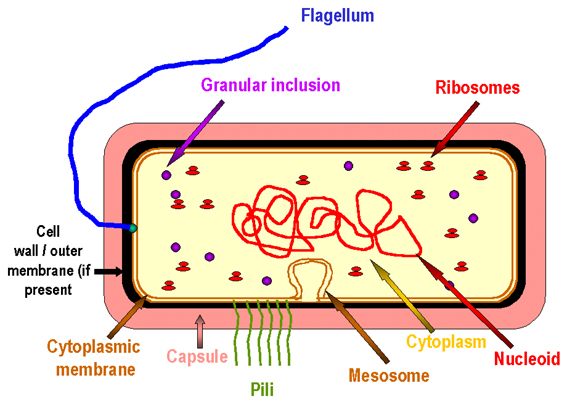 Figure 3
Figure 3
Prototype de cellule bactérienne.
STRUCTURES BACTERIENNES
Malgré leur manque de complexité par rapport aux eucaryotes, un certain nombre de structures des eubactéries doivent être définies. Toutes les bactéries ne possèdent pas tous ces éléments.
Plasmides
Il s’agit d’ADN extra-chromosomique, généralement présent sous forme de plusieurs copies, qui code souvent pour des facteurs de pathogénie et des facteurs de résistance aux antibiotiques. Certaines formes sont aussi impliquées dans la réplication bactérienne.
L’enveloppe cellulaire
Les bactéries peuvent être divisées en deux groupes sur la base de la coloration de Gram ; les bactéries Gram positives restent colorées par le crystal violet après rinçage, pas les Gram négatives. Toutes les bactéries ont une membrane cellulaire où une phosphorylation oxydative a lieu (puisqu’il n’y a pas de mitochondrie). En dehors de la membrane cellulaire on trouve la paroi cellulaire, qui est rigide et protège la cellule de la lyse osmotique. Chez les bactéries à Gram positif, la couche de peptidoglycane de la paroi cellulaire est beaucoup plus épaisse que chez les bactéries à Gram négatif. Les bactéries à Gram négatif ont une membrane externe supplémentaire. Cette membrane externe est la principale barrière de perméabilité chez les bactéries à Gram négatif. L’espace entre la membrane interne et la membrane externe est connu sous le nom d’espace périplasmique. Les bactéries à Gram négatif stockent des enzymes de dégradation dans l’espace périplasmique. Les bactéries à Gram positif n’ont pas d’espace périplasmique ; elles secrètent donc à la place des exoenzymes pour réaliser la digestion extracellulaire. Cette digestion est nécessaire puisque les grosses molécules ne peuvent pas passer directement à travers la membrane externe (si elle est présente) ou la membrane cellulaire.
Formes de bactéries sans paroiQuand les bactéries sont traitées avec 1) des enzymes lytiques pour la paroi cellulaire comme le lysozyme ou 2) des antibiotiques qui interfèrent avec la biosynthèse du peptidoglycane, des bactéries sans paroi sont souvent produites. Généralement ces traitements génèrent des organismes non viables. Les bactéries sans paroi qui ne peuvent pas se répliquer sont dénommées sphéroplastes (quand une membrane externe est présente) ou protoplastes (en absence de membrane externe). Occasionnellement des bactéries sans paroi qui peuvent se répliquer sont générées par ces traitements (formes L).
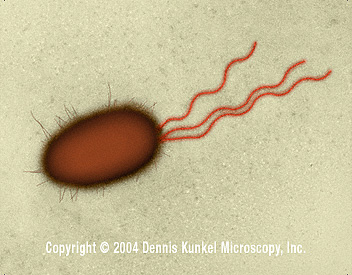 Figure 4
Figure 4
E. coli – bacille procaryote avec de multiples flagelles
(SEM x26,730) ©
Dennis Kunkel Microscopy, Inc.
Used with permission
FlagelleQuelques espèces bactériennes sont mobiles et possèdent des organelles locomoteurs – les flagelles (Figure 4). Celles qui en possèdent sont capables de goûter leur environnement et de répondre à des aliments chimiques spécifiques ou des composés toxiques et de se déplacer vers ou à l’opposer de ces éléments (chimiotaxie). Les flagelles sont intégrés à la membrane cellulaire, s’étendent à travers l’enveloppe cellulaire et sont projetés comme de longs filaments. Les flagelles sont constitués de protéines dont la flagelline. Ils permettent le déplacement de la cellule en tournant à la manière d’une hélice. Les filaments axiaux des spirochètes ont une fonction similaire à celle des flagelles. Des protéines de liaisons de l’espace périplasmique ou de la membrane cellulaire lient des sources de nourriture (telles que les sucres et les acides aminés) conduisant à la méthylation d’autres protéines de la membrane cellulaire, affectant alors le mouvement de la cellule par les flagelles. Des protéines appelées perméases transportent alors ces aliments à travers la membrane cellulaire. L’énergie et les sources de carbone peuvent alors être stockées dans des « granules de stockage » du cytoplasme qui sont constitués de glycogène, polyhydroxybutyrate ou polyphosphate.
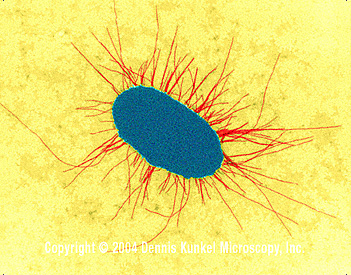 Figure 5
Figure 5 E. coli avec des fimbriae (TEM x17,250). © Dennis Kunkel Microscopy, Inc. Used with permission
Pili (synonym : fimbriae)Le type de pilus (ou le fait qu’ils soient produits ou pas) varie entre les espèces et au sein d’une même espèce. Les pili sont des projections de la cellule de type poils (figure 5). Certains sont impliqués dans la conjugaison sexuelle et d’autres permettent l’adhésion aux surfaces épithéliales de l’hôte lors de l’infection.
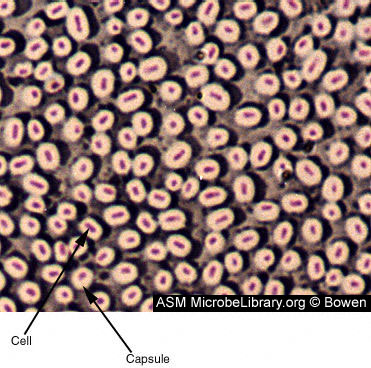 Figure 6 Bactérie en forme de bacille et produisant une
capsule. La capsule est composée de polysaccharides et de polyprotéines. Les
capsules ont un rôle d’adhérence, de virulence, de protection, de capture
des nutriments, et de reconnaissance entre cellules. Les capsules varient en
épaisseur et peuvent facilement être de deux fois le volume de l’organisme.
Lors d’une coloration de capsule, le fond est coloré en bleu-grisâtre et les
cellules sont colorées en rouge. La capsule n’est pas colorée et apparait
comme un halo autour de la cellule. © Judy Bowen
Figure 6 Bactérie en forme de bacille et produisant une
capsule. La capsule est composée de polysaccharides et de polyprotéines. Les
capsules ont un rôle d’adhérence, de virulence, de protection, de capture
des nutriments, et de reconnaissance entre cellules. Les capsules varient en
épaisseur et peuvent facilement être de deux fois le volume de l’organisme.
Lors d’une coloration de capsule, le fond est coloré en bleu-grisâtre et les
cellules sont colorées en rouge. La capsule n’est pas colorée et apparait
comme un halo autour de la cellule. © Judy BowenBuckman Laboratories International, Inc.
Memphis, Tennessee, USA and The MicrobeLibrary
Capsule et couche de slime (Figure 6)
Ce sont des structures entourant l’extérieur de l’enveloppe cellulaire. Lorsque la structure est bien définie, il s’agit de capsule et lorsqu’elle est moins définie il s’agit de couche de slime ou glycocalyx. Elles consistent généralement en polysaccharide cependant chez certains bacilles elles sont composées de polypeptides (acide polyglutamique). Elles sont essentielles pour la viabilité cellulaire et certaines souches au sein d’une espèce vont produire une capsule quand d’autres non. Les capsules des bactéries pathogènes inhibent l’ingestion et la dégradation par les phagocytes. Les capsules sont souvent perdues lors de la culture in vitro.
Endospores (spores)
Ce sont des formes dormantes des cellules bactériennes produites par certaines bactéries en cas de pénuries de nutriments (figure 7) ; la forme activement croissante de la cellule est appelée végétative. La spore est résistante aux conditions difficiles (dont les hautes températures et les solvants organiques). Le cytoplasme de la spore est déshydraté et contient du calcium dipicolinate (acide dipicolonique- figure 8) qui est impliqué dans la résistance de la spore à la chaleur. Les spores sont souvent retrouvées dans les genres Bacillus et Clostridium.
 Figure 7 A
Figure 7 AFigure 7 A : spore de Bacillus cereus (vert) et des cellules ne formant pas de spore (rose). © Ralph Van Dyke Jr. The Des Moines University Osteopathic Medical Center, Des Moines, Iowa USA and The MicrobeLibrary
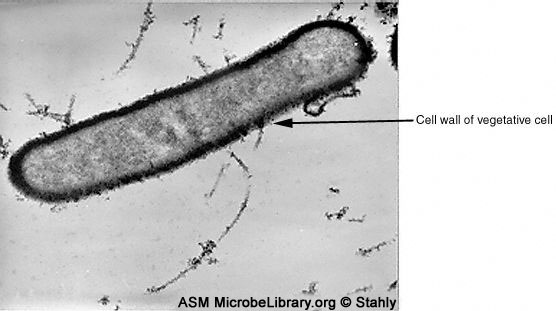 B
B
La sporulation de P.larvae en culture apparait similaire à celle des autres
fabricants d’endospores. La cellule végétative en forme de bacille possède une
épaisse couche de peptidoglycane.
© Donald Stahly, University of Iowa, Iowa City, Iowa
and The MicrobeLibrary
Une spore immature est montrée entourée de la cellule mère (sporangium). Une copie de l’ADN bactérien est enveloppée dans la spore en développement. La couche externe de la spore apparait plus fine et moins dense aux électrons que les spores matures. © Donald Stahly, University of Iowa,
Iowa City, Iowa and The MicrobeLibrary
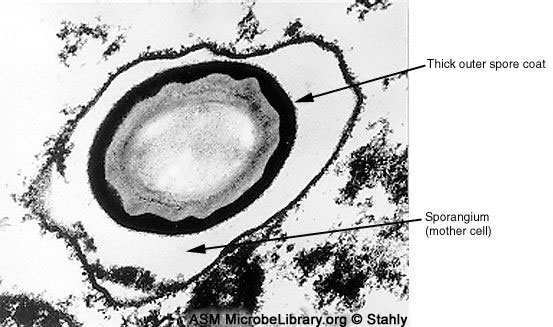 D
D
La couche épaisse de la spore indique que la
différenciation est accomplie, mais l’endospore reste à l’intérieur du
sporangium. Finalement l’endospore est libérée du sporangium. La couche interne
de la spore consiste en un maximum de sept couches distinctes appelées lamelles.
© Donald Stahly, University of Iowa, Iowa City, Iowa
and
The MicrobeLibrary
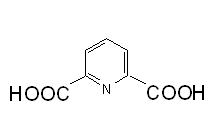 Figure 8. Acide dipicolinique.
Figure 8. Acide dipicolinique.
![]()
Retourner à la section d'bacteriologie de Microbiologie et Immunologie On-line
This page last changed on Tuesday, July 30, 2013
Page maintained by Richard Hunt
Please report any problems to richard.hunt@uscmed.sc.edu