|
x |
x |
|
 |
 |
|
DOENÇAS
INFECCIOSAS |
BACTERIOLOGIA |
IMUNOLOGIA |
MICOLOGIA |
PARASITOLOGIA |
VIROLOGIA |
|
|
VIROLOGIA – CAPÍTULO QUATRO
ESTRATÉGIAS DE REPLICAÇÃO DE VIRUS DE RNA
Dr Margaret Hunt
Professor Emerita
Department of Pathology, Microbiology and Immunology
University of South Carolina School of Medicine
Tradução:
PhD. Myres Hopkins
|
|
En
Español |
|
|
|
NË SHQIPTARE |
Dê a
sua opinião
CONTATO |
|
BUSCA |
E-MAIL
DR MYRES HOPKINS |
|
|

 |
|
Logo image © Jeffrey
Nelson, Rush University, Chicago, Illinois and
The MicrobeLibrary |
|
|
|
OBJETIVOS
Análise descritiva das estratégias replicativas empregadas por vírus de
RNA animais
Identificação de protótipos virais associados com esquemas de replicação
de diferentes tipos de vírus de RNA
 Estrutura do Pólio Tipo 1 Mahoney.
Dados de raios-X de Hogle et al.(Univ. de Harvard), entrada de 2PLV no
PDB, renderizado com GRASP (A.Nicholls, Columbia Univ.). Cortesia do
urtesy of
Dr
Sgro
e do
Institute for Molecular Virology,
Univ. de Wisconsin (usado com permissão)
Estrutura do Pólio Tipo 1 Mahoney.
Dados de raios-X de Hogle et al.(Univ. de Harvard), entrada de 2PLV no
PDB, renderizado com GRASP (A.Nicholls, Columbia Univ.). Cortesia do
urtesy of
Dr
Sgro
e do
Institute for Molecular Virology,
Univ. de Wisconsin (usado com permissão) |
REPLICAÇÃO DE VÍRUS DE RNA - GENERALIDADES
ESTRATÉGIAS
Vírus de RNA
que não têm uma fase de DNA
Vírus que replicam via intermediários de RNA necessitam de uma RNA
polimerase dependente de RNA para replicar seu RNA, mas células animais não
parecem posuir uma enzima adequada. Portanto, este tipo de vírus de RNA
animal necessita de codificar para uma RNA polimerase dependente de RNA.
Nenhuma proteína viral pode ser feita até que o RNA mensageiro viral esteja
disponível; dessa dorma, a natureza do RNA no virion afeta a estratégia do
vírus:
Vírus com fita de RNA positiva
Nesses
vírus, o RNA (genômico) do virion é de mesmo sentido que o RNAm e portanto
funciona como RNAm. Este RNAm pode ser traduzido imediatamente mediante
infecção da célula hospedeira.
Exemplos:
Vírus com fita de RNA negativa
O RNA do
virion tem sentido negativo (complementar ao do RNAm) e precisa portanto
ser copiado no RNAm complementar de sentido positivo para que as proteínas
possam ser feitas. Assim, além de precisar codificar para uma
RNA-polimerase dependente de RNA, esses vírus também precisam empacotá-la
no virion para que ela possa fazer RNAm ao infectar a célula
Exemplos:
Vírus de RNA de fita dupla
O RNA do
virion (genômico) é de fita dupla e portanto não pode funcionar como RNAm;
assim esses vírus também precisam empacotar uma RNA polimerase para fazer
seus RNAm após infecção da célula hospedeira.
Exemplo:
|
VÍRUS DE RNA
QUE NÃO TÊM UMA FASE DE DNA |
|
Genoma |
RNA
polimerase dependente de RNA (=transcritase) no virion |
Infectividade
do RNA |
Evento
inicial na célula |
|
RNA de fita
positiva |
Não |
Infeccioso |
Tradução |
|
RNA de fita
negativa |
Sim |
Não
infeccioso |
Transcrição |
|
Double
-stranded RNA |
Sim |
Não
infeccioso |
Transcrição |
|
RETROVÍRUS |
|
Genoma |
RNA polimerase
dependente de RNA (=transcritase) no virion |
Infectividade
do RNA |
Evento
inicial na célula |
|
RNA de fita
positiva |
Sim |
Não
infeccioso |
Transcrição
reversa |
Vírus de RNA
que copiam seu RNA em DNA
Esses são
os
retrovírus.
Nesse caso, seus virions de RNA, embora de sentido positivo, não funcionam
como RNAm imediatamente após a infecção, visto que ele não é liberado do
capsídeo para o citoplasma. Ao invés disso, ele serve como um template para
a transcritase reversa e é copiado em DNA. A transcritase reversa não está
disponível na célula, e assim esses vírus necessitam codificar para esta
enzima e empacotá-la nos virions.
O PROBLEMA DA
TRADUÇÃO
A maquinária de tradução
ou síntese protêica da célula hospedeira eucariótica em geral usa RNAm
monocistrônicos
e assim existe um problema ao fazer mais de um tipo de proteína a partir de um
único RNAm.
Vírus de RNA têm algumas
soluções para este problema:
- O vírus faz RNAm múltiplos monocistrônicos
- O vírus faz transcritos primários que são processados
pela maquinária de splicing do hospedeiro para fazer mais de um RNA
monocistrônico
- O RNAm viral age como um transcrito monocistrônico.
Um grande polipeptídeo (chamado de poliproteína) é feito, que é então
clivado em proteínas separadas – Assim, um produto de tradução inicial é
processado para produzir proteínas múltiplas. Isso acontece, por exemplo, em
picornavírus
- O RNAm viral tem características especiais que
permitem ribossomos se ligarem internamente, ao invés de (ou também) no
terminal 5’.
TAMANHO DO
GENOMA DE VÍRUS DE RNA
Vírus de RNA tendem a
ter um genoma relativamente pequeno (embora o tamanho do
virion
não seja necessariamente pequeno). Isto é provavelmente porque a falta de
mecanismos de correção de erros de RNA ponha um limite no tamanho dos genomas de
RNA.
A consequência de ter um
genoma pequeno é que vírus de RNA tendem a codificar para apenas algumas
proteínas. Estas incluem a polimerase que pode copiar em um ácido nucleico
complementar (quer seja RNA, ou no caso de retrovírus, DNA) e uma proteína de
ligação viral.
|
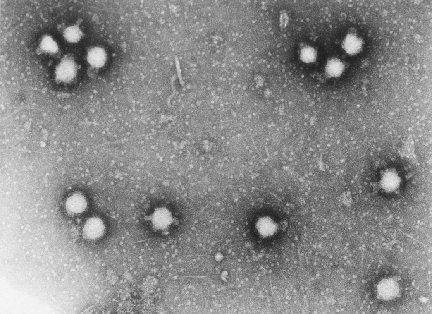
 Figura 1 Vírus da pólio
© J-Y Sgro, Usado com permissão.
De
Virus World
Figura 1 Vírus da pólio
© J-Y Sgro, Usado com permissão.
De
Virus World
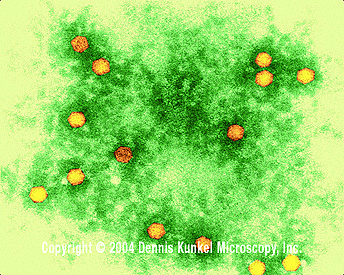
 Figura 2 Vírus da pólio x350,000
©
Dennis Kunkel Microscopy,
Inc.
Usado
com permissão
Figura 2 Vírus da pólio x350,000
©
Dennis Kunkel Microscopy,
Inc.
Usado
com permissão
|
VIRUS DE RNA DE FITA POSITIVA
Exemplos:
·
picornavírus
·
togavírus
·
flavivírus
PICORNAVÍRUS (PICORNAVIRIDAE)
Propriedades
São vírus pequenos
(28nm), nús icosaédricos (figura 1) (pico=muito pequeno). O RNA é de fita
simples, sentido positivo, poliadenilados. Ele funciona como RNAm imediatamente
após a infecção.
Membro protótipo:
poliovírus (figura 1 e 2)
Adsorção e penetração
Uma proteína viral
reconhece um receptor na membrana celular do hospedeiro (é importante para o
tropismo do vírus).
Parece que a ligação ao receptor altera a estrutura do capsídeo de alguma forma,
um canal é formado através da membrana celular e o RNA é liberado no citoplasma.
O RNAm está agora disponível para a tradução.
Síntese de
proteínas virais
O RNA do virion do
poliovirus funciona como um RNAm mas não tem a estrutura cap medilada típica de
RNAm eucarióticos – ele tem uma “pista de pouso do ribossomo” (conhecida como
sítio de entrada interna do ribossomo ou IRES) que habilita os ribossomos a se
ligarem sem ter que reconhecer uma estrutura 5’ cap medilada (figura 3).
Picornavírus frequentemente
interferem no reconhecimento do cap metilado da célula hospedeira. A maioria das
traduções de células hospedeiras são dependentes de cap, assim é inibida muita
síntese protêica do hospedeiro, mas não a síntese protêica viral – uma maneira
pela qual esses vírus podem modificar a célula hospedeira em seu favor.
|
 Figura 3
Figura 3
|
O RNAm é traduzido em um único polipetídeo (poliproteína), que é clivado. As
clivagens ocorrem antes que a tradução seja terminada (ou seja, na cadeia
nascente (= em crescimento) e são realizadas por proteases codificadas pelo
vírus (figura 4). Algumas dessas proteases podem funcionar mesmo enquando são
parte da poliproteína.
|
 Figura 4 Adaptado de Schaechter et al., Mecanismos de Doenças Microbianas, 2nd
Ed.
Figura 4 Adaptado de Schaechter et al., Mecanismos de Doenças Microbianas, 2nd
Ed. |
Os produtos da clivagem
inclui:
Uma RNA polimerase (replicase)
Componentes estruturais das proteases do virion
|
 Figura 5
Figura 5 |
Replicação do
RNA
Nós agora fizemos
proteínas virais novas para apoiar a replicação.
1. A RNA polimerase
viral copia RNA genômico sentido positivo em RNA complementar sentido menos:
Este processo necessita:
VPg (ou precursor
contendo VPg)
RNA polimerase viral (replicase)
Certas proteínas hospedeiras virais
VPg age como um primer
para a síntese de RNA, o que explica por que ele está no terminal 5’ de todas as
moléculas de RNA recentemente sintetizadas.
2. Fitas novas sentido
menos servem de template para novas fitas sentido positivo (figura 5).
Novamente, RNA polimerase e VPg de poliovírus são necessárias. VPg está ligada
aos terminais 5’ das fitas novas sentido positivo (repetindo, elas provavelmente
funcionam como primer).
A nova fita positiva tem três destinos alternativos:
i. Pode servir como um template para mais fitas menos
ii. Pode see empacotada na progênie de virions
iii. Pode ser traduzida em poliproteína (Neste caso VPg é normalmente removida
antes da tradução)
Montagem
Quando é acumulada
uma progênie de RNA sentido mais e proteínas do virion suficiente,
começa a montagem. As partículas são montadas com VPg-RNA dentro e 3
proteínas no capsídeo [VP0,1 e 3]. VP0 é então clivada a VP2 e VP4 à
medida que os virions maturam e esses virions maduros são infecciosos.
Virions são liberados após a lise celular. Excessos de capsídeos são
formados e corpos de inclusão podem ser vistos no citoplasma.
NOTE: O CICLO INTEIRO
OCORRE NO CITOPLASMA
NÃO HÁ DIVISÃO EM EXPRESSÃO GÊNICA INICIAL E TARDIA
|
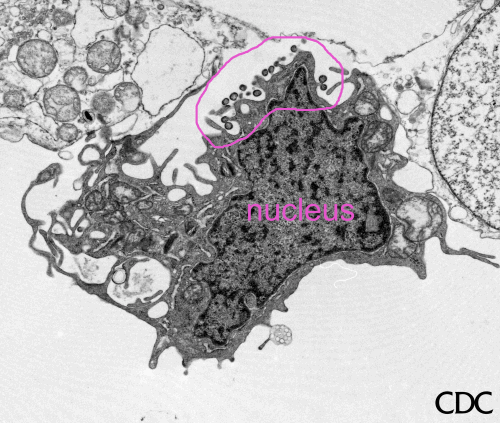
 Figura 6 Rabdovírus em uma Célula Epitelial de Peixe
©
Dennis Kunkel Microscopy, Inc.
Usado
com permissão
Figura 6 Rabdovírus em uma Célula Epitelial de Peixe
©
Dennis Kunkel Microscopy, Inc.
Usado
com permissão
|
VÍRUS NÃO SEGMENTADOS DE FITA NEGATIVA
Exemplos:
|
 Figura 7
Figura 7
Estrutura de um rabdovírus típico
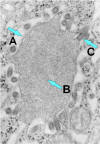 Figura 7b
Figura 7b
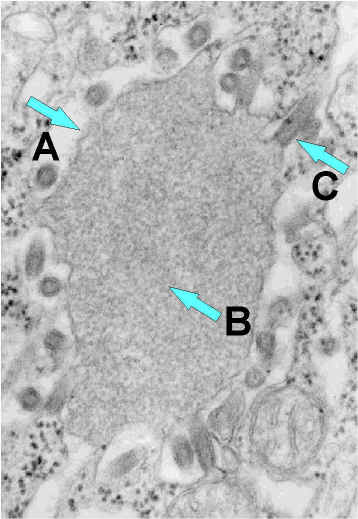
Vírus da
raiva brotando de um corpo de inclusão (corpúsculo negro) para o retículo
endoplasmático em uma célula nervosa. A. Corpo negro. B. Notar o RNP
abundante na inclusão. C. Vírus da raiva em brotamento. CDC |
RABDOVíRUS
(RABDOVIRIDAE)
Exemplo:
Vírus da raiva.
O membro mais intensivamente estudado é o vírus da estomatite vesicular.
RNA:
·
é de fita simples
·
é de sentido negativo
(menos)
·
codifica para 5 proteínas
Ligação,
penetração e desnudamento
O vírus adsorve à superfície celular.
A glicoproteína G é a proteína de ligação (figura 7) que se liga a um receptor
na superfície da célula hospedeira.
O vírus ligado é captado por endocitose.
A membrana do vírus se fusiona com a membrana do endossomo (o pH ácido do
endossomo é importante porque a proteína G necessita ser exposta a pH ácido para
poder facilitar a fusão.
Como resultado da fusão da membrana viral com a do endossomo, o nucleocapsídeo é
liberado no citoplasma.
Transcrição
'Transcrição' é usado neste contexto para se referir à síntese de RNAm.
O desnudamento completo do nucleocapsídeo não é necessário para a transcrição –
a RNA polimerase do virion pode copiar o RNA do virion quanto ele está na sua
forma nucleocapsídica. Isso é uma vantagem no sentido de que o RNA genômico fica
um pouco mais protegido de ribonucleases.
Há um RNAm monocistrônico para cada uma das cinco proteínas codificadas pelo
vírus (figura 8). Os RNAm recebem o 5’ cap, é metilado e poliadenilado. Visto
que é um vírus de RNA de sentido negativo citopasmático, as enzimas da síntese
de RNA e modificação estão empacotadas no virion.
Tradução
RNAs mensageiros são traduzidos nos ribossomos hospedeiros e todas as cinco
proteínas virais são feitas ao mesmo tempo. Não há distinção entre funções
iniciais e tardias.
|
 Figura 8
Figura 8 |
Replicação do
RNAA
replicação do RNA é o processo pelo qual novas cópias do comprimento do genoma
de RNA são feitas (figura 8).
A replicação do RNA ocorre no citoplasma e é realizada pela RNA polimerase
viral.
A fita mais é coberta em todo o seu comprimento por proteínas do nucleocapsídeo
à medida que é feita (RNAm não são cobertos com esta proteína, o que poderia
interferir na maquinária de tradução de proteínas hospedeiras).
A nova fita positiva é
copiada em uma fita menos completa, que é também coberta por proteínas do
nucleocapsídeo à medida que é feita. (Note: visto que a RNA polimerase viral
sintetiza RNAm (transcrição) e toda a extensão do RNA (replicação), ela é
às vezes chamada de transcritase ou replicase, sendo que tais nomes focalizam
diferentes aspectos da atividade da polimerase).
A fita negativa nova
pode:
i. ser usada como
template para a síntese de mais fitas mais inteiras
ii. ser usadas como templates para a síntese de mais RNAm
iii. ser empacotadas em virions
|
 Figura 9
Figura 9
|
Montagem
O vírus consiste de
dois "módulos" – o envelope e o nucleocapsídeo:
Envelope
Proteínas transmembrana são feitas nos ribossomos ligados ao retículo
endoplasmático. Elas são inseridas no retículo endoplasmático à medida que
são feitas, glicosiladas no retículo endoplasmáttico e passam através do
corpúsculo de Golgi, onde modificações substanciais das cadeias de
carboidratos ocorrem. Elas são então transportadas em vesículas para a
membrana celular apropriada; no caso do vírus da estomatite vesicular, esta
é a membrana plasmática (figura 9).
|
 Figura 10
Figura 10
|
Nucleocapsidio
A síntese do nucleocapsídeo foi descrita
acima. O complexo da RNA polimerase viral associa com os nucleocapsídeos
a medida que eles são formados.
Nucleocapsídeos
brotam através de áreas modificadas da membrana que contém proteínas G e
M (figura 10). A proteína M (matriz) está envolvida na montagem – ela
interage com regiões de G na membrana e com nucleocapsídeos.
NOTE:
·
O CICLO DE VIDA
INTEIRO OCORRE NO CITOPLASMA
·
A RNA POLIMERASE E ENZIMAS DE
MODIFICAÇÃO DO RNA SÃO CODIFICADAS PELO VÍRUS E ESTÃO PRESENTES NO
VIRION
NÃO HÁ DIVISÃO
ENTRE INICIAL E TARDIO
|
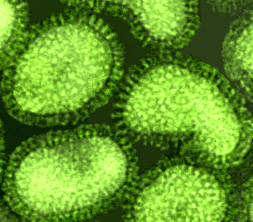
 Figura 11 Paramixovírus ©
Dr
Linda Stannard,
Universidade de Cape Town, África do Sul (usado com permissão)
Figura 11 Paramixovírus ©
Dr
Linda Stannard,
Universidade de Cape Town, África do Sul (usado com permissão)
|
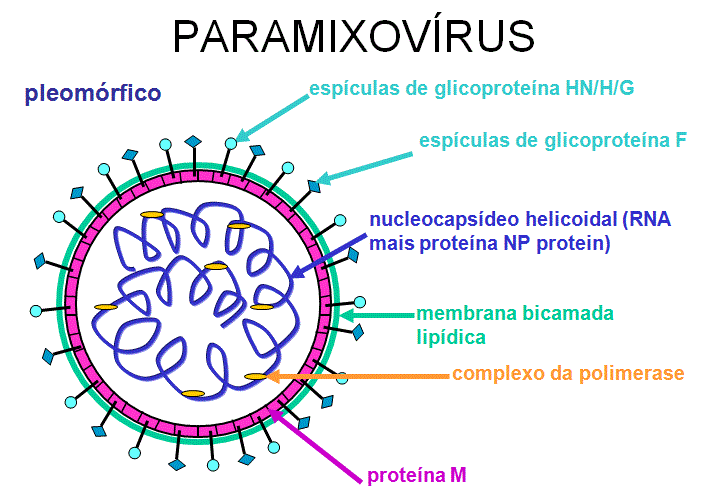
PARAMIXOVÍRUS
(PARAMIXOVIRIDAE)
Paramixovírus
(figura 11) são pleomórficos, ou seja: há muitas formas morfológicas do
vírus em uma população. Eles têm RNA de sentido negativo, não segmentado e
um nucleocapsídeo helicoidal (figura 12). Eles são envelopados, isto é, eles
são arrodeados por uma membrana derivada de uma célula hospedeira.
O envelope contém duas glicoproteínas codificadas pelo vírus: A proteína F e
a proteína de ligação
-
A
proteína F tem atividade de fusão
-
A
proteína de ligação se liga a receptores na célula hospedeira
Esta proteína pode ter:
Atividade
hemaglutinante
e atividade de
neuraminidase
(proteína HN) ou atividade hemaglutinante sozinha (proteína H) ou nenhuma
(proteína G).
|
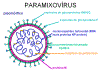 Figura
12 Estrutura de um paramixovírus típico
Figura
12 Estrutura de um paramixovírus típico |
|
GLICOPROTEÍNAS DE SUPERFÍCIE DA FAMÍLIA DOS PARAMIXOVÍRUS |
|
GÊNERO
|
GLICOPROTEÍNA
|
MEMBROS TÍPICOS
|
|
FAMÍLIA
DOS PARAMYXOVIRUS |
|
Paramixovírus
|
HN, F
|
HPIV 1
HPIV 3
|
|
Rubulavírus |
HN, F |
HPIV 2
HPIV 4
vírus da
caxumba |
|
Morbilivírus
|
H, F
|
virus do sarampo
|
|
FAMÍLIA
DOS PNEUMOVÍRUS |
|
Pneumovírus
|
G, F
|
vírus respiratório sincicia
|
|
Metapneumovírus |
G, F |
metapneumovírus |
Hemaglutinação
Hemaglutinação é fácil de testar em um laboratório clínico e é usada
no diagnóstico.
A
hemaglutinação envolve a aglutinação de células vermelhas do sangue
e se baseia na capacidade de um vírus de se ligar a receptores nas
células vermelhas sanguíneas. Visto que vírus têm múltiplas
proteínas de ligação por virion, elas podem se ligar a mais de uma
célula vermelha e assim eles podem servir para ligar essas células
vermelhas em uma malha. Vírus inativados ainda podem hemaglutinar se
as suas proteínas de ligação estiverem intactas.
Se alguma
pessoa tem anticorpos para uma hemaglutinina viral, os anticorpos
irão se ligar à proteína de ligação e impedir sua ligação às células
vermelhas do sangue. O soro daquela pessoa irá inibir a reação de
aglutinação pelo vírus para o qual ela tem anticorpos – mas não por
outros vírus hemaglutinadores. Isso pode ser usado para determinar a
qual o vírus hemaglutinador a pessoa foi exposta.
Hemadsorção
Durante a infecção, a proteína de ligação
viral será inserida na membrana plasmática da célula infectada. Se a
proteína de ligação viral puder se ligar a células vermelhas do
sangue, a célula infectada irá se ligar a células vermelhas do
sangue porque elas têm a proteína de ligação viral na sua superfície
– isso é chamado de hemadsorção. No laboratório clínico, isso pode
fazer com que células infectadas por vírus sejam detectadas em um
estágio inicial da infecção, e pode permitir a detecção de vírus que
não danificam a célula de maneira visível.
|
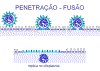 Figura 13
Figura 13 |
Adsorção
e penetração
A proteína H(N)/G
reconhece receptores na superfície celular.
A proteína F
facilita a fusão entre membranas em pH fisiológico, e embora paramixovírus
possam ser captados por endocitose, eles também frequentemente entram na
célula por fusão direta com a membrana plasmática (figura 13).
Devido à proteína F funcionar em pH fisiológico, isso
pode resultar na formação de um
sincício sendo
formado em infecções por paramixovírus (ver discussão sobre consequências da
fusão em pH fisiológico em
vírus de DNA – estratégias de replicação
– vírus do herpes).
|
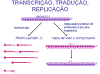 Figura 14
Figura 14 |
Transcrição, tradução e replicação do RNA
Eventos no interior
da célula são muito similares ao rabdovírus (figura 14):
·
A
multiplicação viral ocorre no citoplasma.
·
A RNA polimerase viral usa o nucleocapsídeo
como template.
·
A RNA
polimerase não necessita um nucleocapsídeo totalmente coberto.
·
RNAm
virais são transcritos; eles recebem o cap, são metilados e poliadenilados.
·
Visto que é um vírus de RNA de fita
negativa, a RNA polimerase e enzimas de modificação do RNA são empacotadas
no virion.
·
Os RNAm
virais são traduzidos para produzir proteínas virais.
·
Não há
distinção entre funções iniciais e tardias na expressão gênica.
Replicação de RNA
viral envolve a síntese de todo o comprimento da fita mais. Ela é usada como
template para uma fita de todo o comprimento sendo menos. Ambas as fitas de
comprimento completo são cobertas com proteína do nucleocapsídeo à medida
que elas são feitas (figura 14).
Fitas novas menos de
comprimento completo podem servir como templates para a replicação, ou
templates para a transcrição, ou elas podem ser empacotadas em novos
virions.
|
 Figura 15
Figura 15
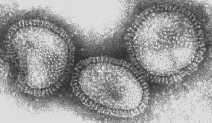
 Figura 16 Ortmixovírus (Influenza A)
© Dr Linda Stannard, Universidade da Cidade do Cabo, África do Sul
Figura 16 Ortmixovírus (Influenza A)
© Dr Linda Stannard, Universidade da Cidade do Cabo, África do Sul
|
Montagem
As glicoproteínas
virais (proteínas de ligação e proteína F de fusão) são traduzidas como
proteínas transmembrana e transportadas para a membrana citoplasmática.
A proteína M (matriz) habilita nucleocapsídeos a interagir com as regiões da
membrana plasmática que têm as glicoproteínas inseridas.
O vírus brota através da membrana.
Papel da
neuraminidase
Nos paramixovírus
que as possuem, a neuraminidase pode facilitar a liberação. Nesses vírus, o
ácido siálico aparenta ser uma parte importante do receptor. A neuraminidase
remove ácido siálico (ácido neuramínico) da superfície da célula. Assim,
como o ácido siálico vai estar bastantee removido da superfície da célula e
da progênie de vírions, não haverá receptores funcionais. Assim, a progênie
de virions não irá se ligar um ao outro ou à célula de onde eles acabaram de
brotar (ou a qualquer outra célula infectada). Eles vão portanto ser capazes
de se difundir para longe até encontrarem uma célula não infectada.
A neuraminidase pode
também ajudar durante a infecção pois se o vírus se liga a resíduos do ácido
siálico no muco, ele não será capaz de se ligar a um receptor de uma célula
e infectar esta célula. Mas se o ácido siálico no muco for eventualmente
destruído, o vírus será libertado e pode então atingir o receptor na
superfície da célula.
Ativação
da proteína F de fusão
A proteína F
necessita ser clivada para poder funcionar facilitando a fusão quando o
vírus se liga a outra célula (figura 15). Este é um evento tardio na
maturação.
|
Algumas
diferenças entre rabdovírus e paramixovírus |
| |
Rabdovírus |
Paramixovírus |
|
Forma |
projétil
baciliforme |
redondo
pleomórfico |
|
Glicoproteínas |
Uma (tem
atividades de ligação e de fusão) |
Duas (uma
de ligação e outra de fusão) |
|
pH de
fusão |
ácido |
neutro
fisiológico |
|
|
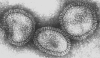 Figura 17 Ortomixovírus (Influenza A)
© Dr Linda Stannard, Universidade da Cidade do Cabo, África do Sul
Figura 17 Ortomixovírus (Influenza A)
© Dr Linda Stannard, Universidade da Cidade do Cabo, África do Sul
 Figura 18 Buniavírus
Figura 18 Buniavírus
Das bases de dados do ICTV
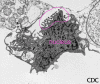 Figura 19b
Células de cultura de tecido vero E6 infectadas com arenavirus. A imagem
mostra partículas virais extracelulares brotando da superfície da
célula. Aumento aprox. 12,000 X.Cortesia
da imagem de Cynthia Goldsmith, MS, Atividade Patológica em Doenças
Infecciosas, DVRD, NCID, CDC
Figura 19b
Células de cultura de tecido vero E6 infectadas com arenavirus. A imagem
mostra partículas virais extracelulares brotando da superfície da
célula. Aumento aprox. 12,000 X.Cortesia
da imagem de Cynthia Goldsmith, MS, Atividade Patológica em Doenças
Infecciosas, DVRD, NCID, CDC |
VÍRUS SEGMENTADOS DE FITA NEGATIVA
Exemplos:
·
Ortomixovírus (figura 16 e 17)
·
Buniavírus (incluem o gênero dos Hantavírus) (figura 18)
·
Arenavírus (figura
19b)
ORTOMIXOVÍRUS (ORTOMIXOVIRIDAE)
Ha três grupos de
vírus da
influenza:
A, B e C. O vírus da influenza A é mais intensivamente estudado e
influenza A e B são os mais importantes na doença humana.
Vírus da influenza
são virions pleomórficos (isto é, eles variam de forma). Eles têm sentido
negativo, fita de RNA simpes e um genoma de RNA que é SEGMENTADO. Existem 8
segmentos na influenza A. O nucleocapsídeo é helicoidal (figura 19). Os
virions contèm RNA polimerase empacotada dentro da partícula viral
Esse vírus são envelopados e têm duas membranas de glicoproteínas (figura
19):
- HA –
hemaglutinina - Esta é a proteína de ligação e fusão
-
NA - neuraminidase
- Esta é importante na liberação. Ela remove o ácido
siálico de proteínas do vírus e da célula hospedeira
|
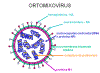 Figura 19 Estrutura de um ortomixovírus típico
Figura 19 Estrutura de um ortomixovírus típico |
Adsorção e
penetração
O vírus adsorve a receptores na superfície da célula e é internalizado
por endocitose. Em pH ácido de um endossomo, HA realiza uma mudança
conformacional e ocorre a fusão.
Os
nucleocapsídeos são liberados no citoplasma.
|
 Figura 20
Figura 20 |
Transcrição, tradução e replicação
Nucleocapsídeos são
transportados para o núcleo. A síntese de RNAm e replicação do RNA viral
ocorre no núcleo. Isso é muito incomum para um vírus de RNA. O vírus
da influenza tem um mecanismo não usual de adquirir um terminal 5’ cap
metilado para o seu RNAm.
Uma endonuclease
viral (que é empacotada no vírus da influenza) desliza do terminal 5’ de um
RNAm hospedeiro capeado e metilado cerca de 13-15 bases a partir do terminal
5’ e usa isso como um primer para a síntese do RNAm viral (figura 20) –
portanto, todos os RNAm do vírus da influenza têm um pequeno trecho no
terminal 5’ que é derivado do RNAm hospedeiro.
A RNA polimerase
viral (transcritase) se extende do primer e copia o template em RNAm
complementar sentido positivo e adiciona uma cauda poli-A. A transcrição
resulta em 8 transcritos primários, um transcrito por segmento. Alguns
segmentos originam transcritos primários que podem ser submetidos a splicing
alternativo (visto que a síntese do RNA do vírus da influenza ocorre no
núcleo, ele tem acesso à maquinária de splicing), cada um originando dois
transcritos alternativos. Por exemplo, o seggmento M origina dois RNAm
alternativos. Estes codificam para a proteína M1 e a proteína M2. Assim, um
único segmento pode codificar para mais de uma proteína pois o vírus tem
acesso à maquinária de splicing. Os RNAm são traduzidos no citoplasma.
Proteínas transmembrana se movem para a membrana plasmática enquanto
proteínas necessárias para a replicação do RNA são transportadas para o
núcleo.
|
| |
Replicação do
RNA
A replicação do
RNA ocorre no núcleo usando uma enzima codificada pelo vírus (esta pode ser a
mesma que a da RNA polimerase envolvida na transcrição de RNAm, ou uma versão
modificada). Um cópia completa exata complementar do RNA do virion é feita –
este RNA sentido positivo é provavelmente coberto com proteína do nucleocapsídeo
enquanto é feito. Um RNA fita positiva completo é então usado como template para
a síntese de uma fita completa sentido negativo; novamente a nova fita menos é
provavelmente coberta com proteína do nucleocapsídeo enquanto ele é feito. Novas
fitas menos podem ser usadas como templates para replicação, síntese de RNAm, ou
empacotadas.
Montagem
Isso ocorre na membrana plasmática. Nucleocapsídeos são transportados para fora
do núcleo wnquanto proteínas do envelope são transportadas via corpúsculo de
Golgi para a membra citoplasmática. A proteína M1 interage com o nucleocapsídeo
e com uma região modificada da membrana plasmática que contém as glicoproteínas
HA e NA. Os virus então brotam através da membrana celular hospedeira.
NOTE:
·
HA precisa ser clivada para poder promover a
fusão. A clivagem ocorre à medida que o vírus deixa a célula ou no fluido
extracelular. A necessidade de clivagem afeta quais os tecidos podem produzir
infecções virais. A proteína clivada precisa então realizar uma mudança
conformacional, normalmente causada pela exposição a um ambiente endossomal
ácido quando ela infecta a próxima célula, para poder fazer a fusão.
·
NA
provavelmente ajuda o vírus a deixar a célula removendo o ácido siálico dos
receptores. NA também pode ajudar o vírus a penetrar o muco e atingir células
epiteliais do trato respiratório ao permitir que ela dissocie dos receptores
contendo ácido siálico no muco pela destruição dos mesmos. A neuraminidase não
impede o vírus de infectar novas células porque a endocitose é presumivelmente
mais rápida do que a remoção do receptor.
Há similaridades e diferenças entre a família dos Paramixovírus e dos
Ortomixovírus. Membros de ambas as famílias são envelopados, ambos contêm fita
simples de RNA sentido negativo, têm nucleocapsídeos helicoidais. Entretanto, as
duas famílias são muito diferentes. NÃO há relação imunológica entre as duas
famílias.
|
| |
|
PROPRIEDADE
|
PARAMYXOVIRIDAE
|
ORTHOMYXOVIRIDAE
|
|
Genoma
|
não segmentado
|
segmentado
|
|
Síntese de RNA
|
citoplasmático
|
nuclear
|
|
Necessidade de um primer de RNAm
|
não
|
sim
|
|
Hemaglutinina,neuraminidase
|
se ambos, parte da mesma proteína (HN)
|
Influenza A e B têm ambos mas em 2 proteínas diferentes (HA e NA)
|
|
Formação sincicial
|
sim (F funciona em pH fisiológico normal)
|
não (HA funciona em pH ácido)
|
|
 Figura 21
Virion de Reovírus de mamífero
Figura 21
Virion de Reovírus de mamífero
Os dados de cryoEM foram do Laboratório de Tim Baker, Universidade de
Purdue. Os vídeos foram criados por Stephan Spencer. Copyright 1999 Dr Tim
Baker e Stepthen M Spencer. De Dr J-Y Sgro's
Virusworld |
VÍRUS DE RNA DE FITA DUPLA
FAMÍLIA
REOVÍRUS (REOVIRIDAE)
A família dos
Reovírus incluem:
·
os
membros do gênero reovírus
·
os membros
do gênero rotavírus
·
os membros
do gênero orbirírus
·
vírus
colorado da febre do piolho
|
 Figura 22 Estrutura de um reovírus típico
Adaptado de Joklik et al. Zinsser Microbiology 20th Ed.
Figura 22 Estrutura de um reovírus típico
Adaptado de Joklik et al. Zinsser Microbiology 20th Ed. |
Reovirus têm
simetria icosaédrica e um capsídeo multicamada (capsídeo interior e
exterior) (figura 22).
O RNA é de dupla fita. Extistem 10-12 segmentos( dependendo do gênero da
família de Reovírus da qual pertence o vírus) (figura 22).
Há algumas diferenças significantes no ciclo de vida de membros da família
dos reovírus e dos rotavírus. Devido às suas importâncias clínicas em
humanos, nós focalizamos em rotavírus.
|
 Figura 23 Rotavírus (Uma partícula de capsídeo dupla (esquerda), e um
capsídeo interior simples (direita))
Copyright
Dr Linda
Stannard,
Universidade da Cidade do Cabo, África do Sul
Figura 23 Rotavírus (Uma partícula de capsídeo dupla (esquerda), e um
capsídeo interior simples (direita))
Copyright
Dr Linda
Stannard,
Universidade da Cidade do Cabo, África do Sul |
ROTAVIRUS
(rota = roda (para a
aparência de virions no microscópio eletrônico) (figura 23)
Adsorção,
penetração e desnudamento
Ainda não está claro o que exatamente acontece in vivo durante a
entrada dos
rotavírus
na célula. Parece haver uma necessidade de uma protease para remover um
pouco da camada externa do capsídeo e para gerar uma “partícula
intermediária sub-viral " (ISVP) para que o vírus possa entrar no
citoplasma. In vivo, as ISVPs são provavelmente geradas por digestão
por proteasee no trato GI. Uma proteína de ligação viral é então exposta no
ISVP, provavelmente nos vértices, e se liga a receptores da célula
hospedeira. O ISVP ativado entra no citoplasma diretamente ou via
endocitose. No citoplasma, o RNA do virion é copiado pela RNA polimerase
viral enquanto ainda em um nucleocapsídeo que tem menos proteínas associadas
com ele do que estão associadas com o ISVP ou com o virion.
Transcrição e tradução
RNA de fita dupla não funciona como um RNAm e porisso a etapa inicial é
fazer RNAm (transcrição).
Os RNAm são feitos por RNA polimerase codificada pelo vírus empacotada no
virion. O RNA é capeado e metilado pelas enzimas empacotadas no virion. Ele
é então extruído dos vértices do capsídeo.
|
 Figura 24
Figura 24 |
Os RNAm são
traduzidos e as proteínas virais resultantes montam para formar um
capsídeo imaturo. Os RNAm são empacotados em capsídeo imaturo e são
então copiados dentro do capsídeo para formar RNAs de fita dupla (não se
sabe como o vírus garante que cada partícula obtém uma cópia dos 11 RNAm
diferentes) (figura 24). Mais RNAs são agora feitos pelos recentemente
formados capsídeos imaturos.
Montagem
Mais
proteínas são feitas e eventualmente os capsídeos imaturos brotam para o
lumen do retículo endoplasmático. Ao fazer isso, eles adquirem um
envelope temporário que é perdido à medida que ele matura. Esta é uma
característica muito estranha dos rotavírus.
Liberação
Isso provavelmente ocorre via lise celular.
NOTE: O CICLO INTEIRO DE REPLICAÇÃO OCORRE NO CITOPLASMA
|
|
|
 Voltar
à Seção Virologia do Microbiologia e Imunologia Online
Voltar
à Seção Virologia do Microbiologia e Imunologia Online
This page last changed on
Monday, February 08, 2021
Página mantida por
Richard Hunt
|
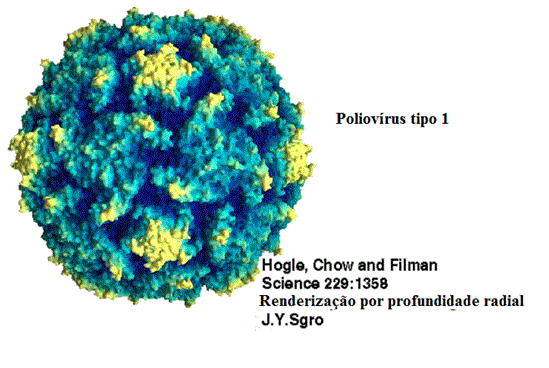 Figura 1 Vírus da pólio
© J-Y Sgro, Usado com permissão.
De
Virus World
Figura 1 Vírus da pólio
© J-Y Sgro, Usado com permissão.
De
Virus World
 Figura 3
Figura 3
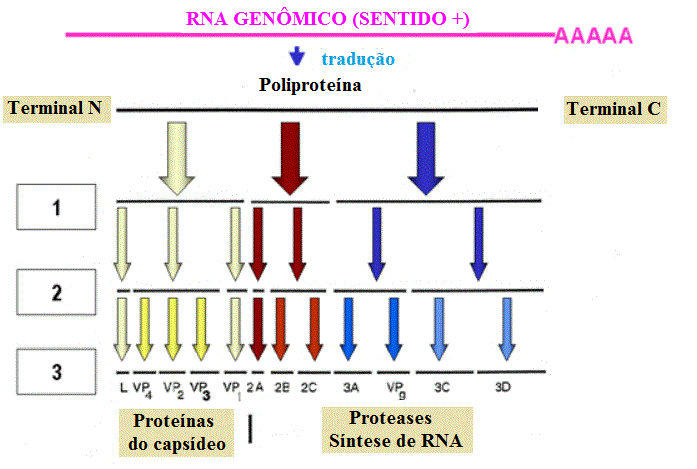 Figura 4 Adaptado de Schaechter et al., Mecanismos de Doenças Microbianas, 2nd
Ed.
Figura 4 Adaptado de Schaechter et al., Mecanismos de Doenças Microbianas, 2nd
Ed.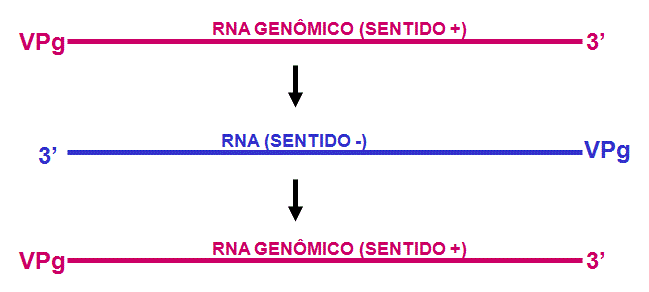 Figura 5
Figura 5 Figura 6 Rabdovírus em uma Célula Epitelial de Peixe
©
Dennis Kunkel Microscopy, Inc.
Usado
com permissão
Figura 6 Rabdovírus em uma Célula Epitelial de Peixe
©
Dennis Kunkel Microscopy, Inc.
Usado
com permissão
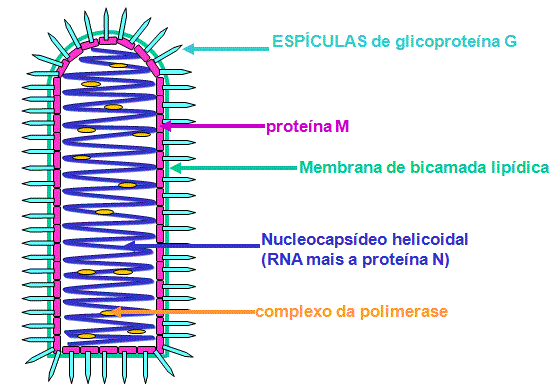 Figura 7
Figura 7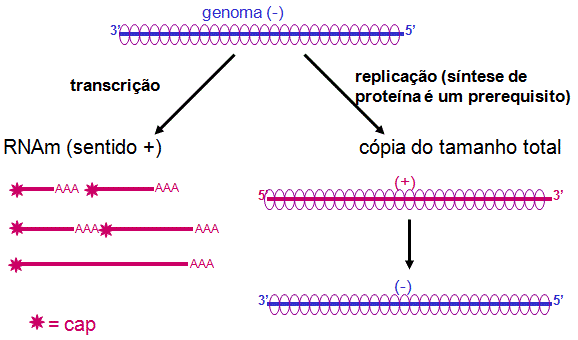 Figura 8
Figura 8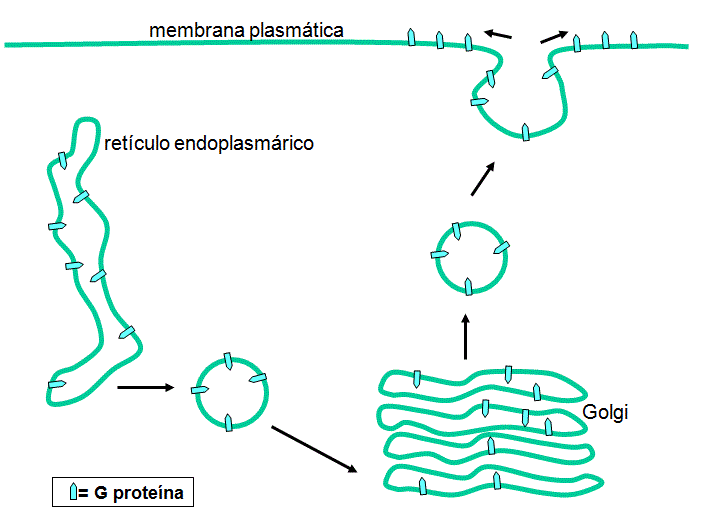 Figura 9
Figura 9
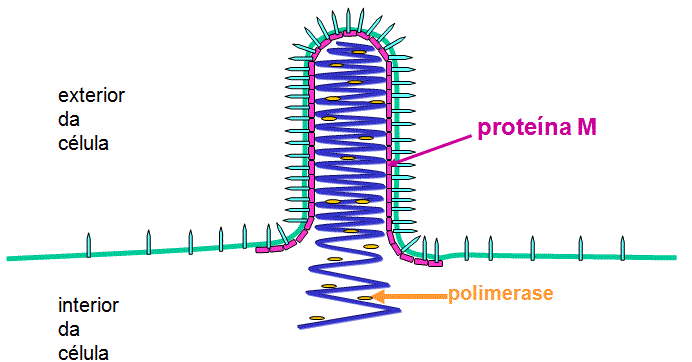 Figura 10
Figura 10
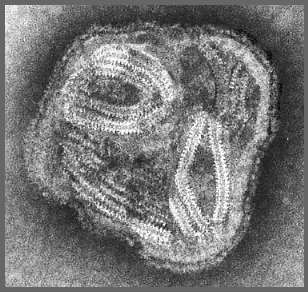 Figura 11 Paramixovírus ©
Dr
Linda Stannard,
Universidade de Cape Town, África do Sul (usado com permissão)
Figura 11 Paramixovírus ©
Dr
Linda Stannard,
Universidade de Cape Town, África do Sul (usado com permissão)
 Figura
12 Estrutura de um paramixovírus típico
Figura
12 Estrutura de um paramixovírus típico Figura 13
Figura 13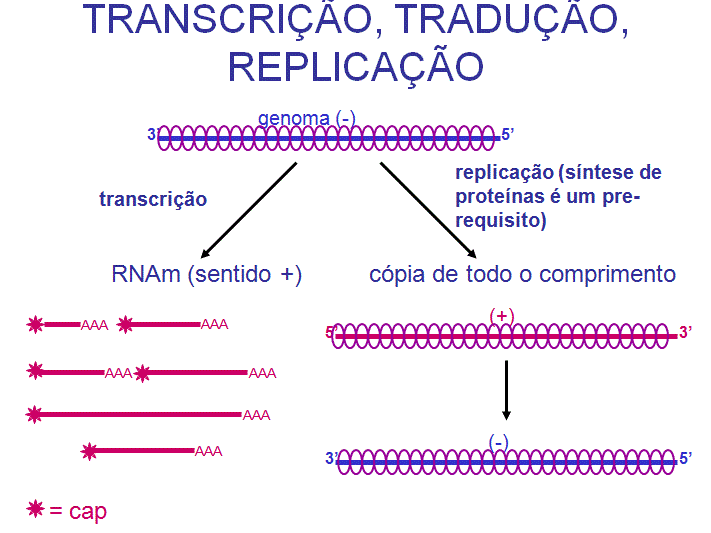 Figura 14
Figura 14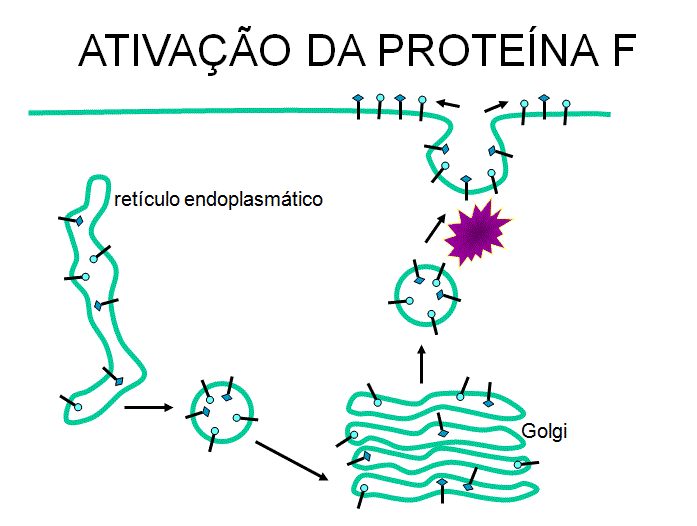 Figura 15
Figura 15
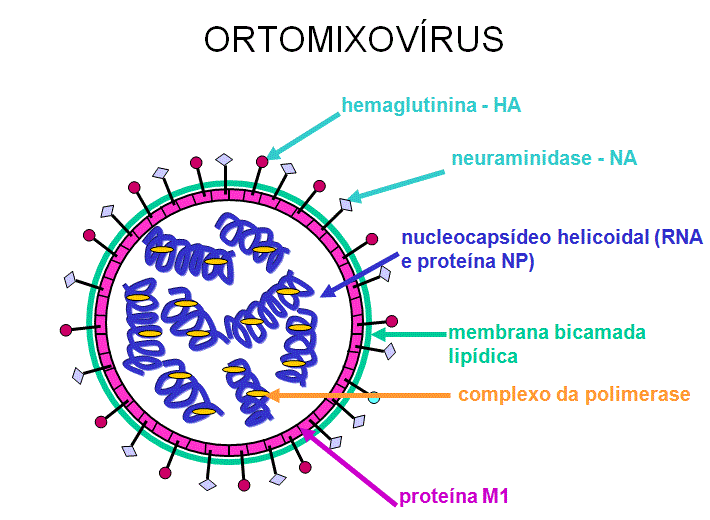 Figura 19 Estrutura de um ortomixovírus típico
Figura 19 Estrutura de um ortomixovírus típico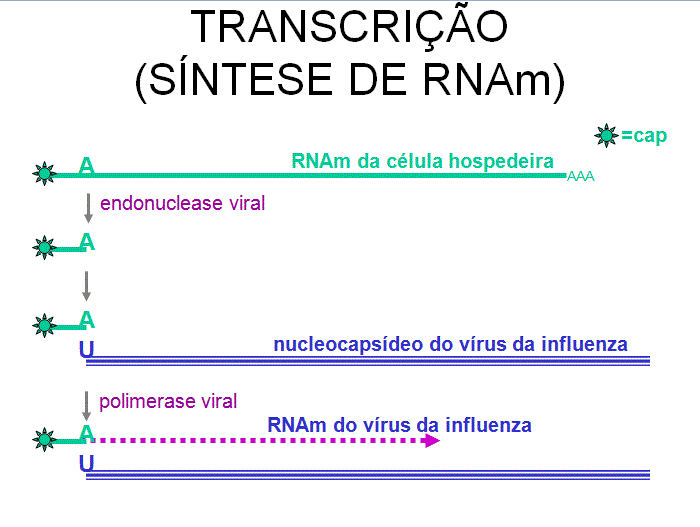 Figura 20
Figura 20 Figura 21
Virion de Reovírus de mamífero
Figura 21
Virion de Reovírus de mamífero 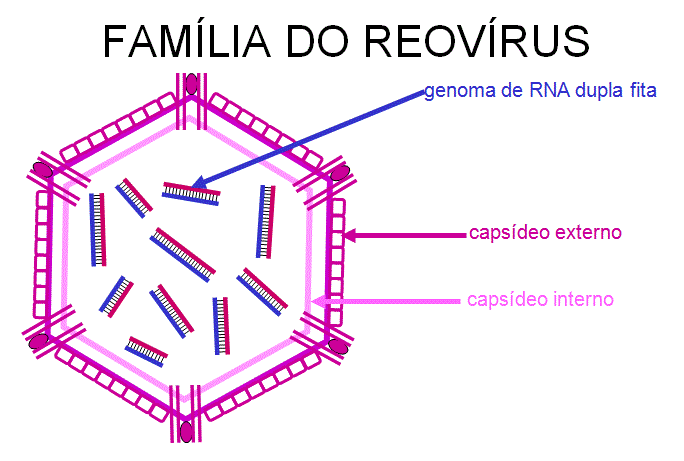 Figura 22 Estrutura de um reovírus típico
Adaptado de Joklik et al. Zinsser Microbiology 20th Ed.
Figura 22 Estrutura de um reovírus típico
Adaptado de Joklik et al. Zinsser Microbiology 20th Ed.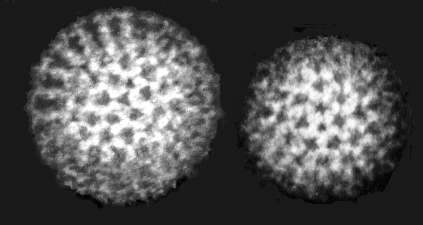 Figura 23 Rotavírus (Uma partícula de capsídeo dupla (esquerda), e um
capsídeo interior simples (direita))
Copyright
Dr Linda
Stannard,
Universidade da Cidade do Cabo, África do Sul
Figura 23 Rotavírus (Uma partícula de capsídeo dupla (esquerda), e um
capsídeo interior simples (direita))
Copyright
Dr Linda
Stannard,
Universidade da Cidade do Cabo, África do Sul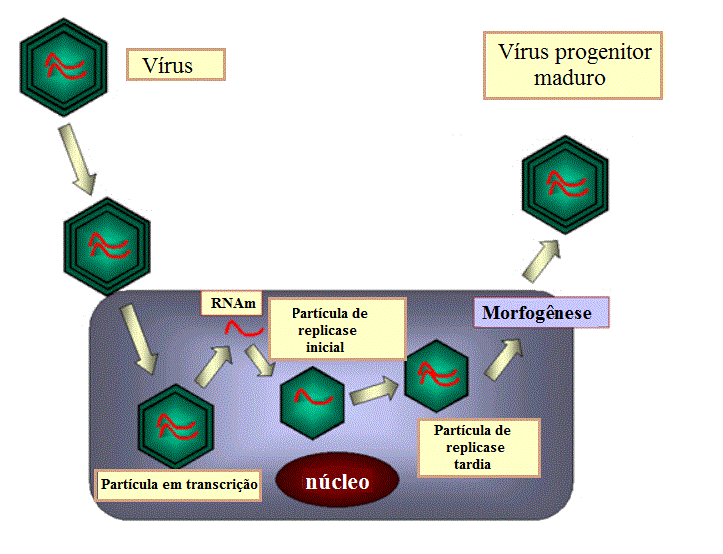 Figura 24
Figura 24