|
x |
x |
|
 |
 |
|
INFECTIOUS
DISEASE |
BACTERIOLOGIE |
IMMUNOLOGIE |
MYCOLOGY |
PARASITOLOGY |
VIROLOGIE |
|
VIDEO LECTURE |
BACTERIOLOGIE – CHAPITRE HUIT
ECHANGE D’INFORMATION GENETIQUE
Gene Mayer, PhD
University of South Carolina School of Medicine
Columbia SC
USA
Emilie Camberlein, PhD
Maître de conférence en Biochimie
Universite de Nantes
Faculte des Sciences et des Techniques
|
|
ANGLAIS |
|
ESPAGNOL |
|
|
Let us know what you think
FEEDBACK |
|
SEARCH |
|
|
|

 |
|
Logo image © Jeffrey
Nelson, Rush University, Chicago, Illinois and
The MicrobeLibrary |
|
|
|
|
INTRODUCTION
Dans les populations bactériennes des mutations apparaissent en permanence à
cause d’erreurs ayant lieu durant la réplication. S’il existe un quelconque
avantage sélectif pour une mutation particulière (ex : résistance aux
antibiotiques), le mutant va alors rapidement devenir le composant majeur de
la population à cause du taux de croissance rapide des bactéries. De plus,
comme les bactéries sont des organismes diploïdes, même les mutations qui
pourraient normalement être récessives seront exprimées. Ainsi, les
mutations des populations bactériennes peuvent poser un problème dans le
traitement des infections bactériennes. Non seulement les mutations sont un
problème mais les bactéries possèdent des mécanismes permettant le transfert
de gènes à d’autres bactéries. Ainsi, une mutation apparaissant dans une
cellule peut être transmise aux autres cellules.
Le transfert de gène chez les bactéries est unidirectionnel d’une cellule
donneuse à une cellule receveuse et le donneur donne généralement seulement
une petite partie de son ADN au receveur. Ainsi, non pas des zygotes
complets mais plutôt zygotes partiels (mérozygotes) sont formés.
Les gènes bactériens sont généralement transférés à des membres de la même
espèce mais occasionnellement le transfert vers d’autres espèces peut avoir
lieu.
|
MOTS CLES
Mérozygote
Transformation
Compétence
Recombinaison homologue
Transduction
Transduction généralisée
Transduction spécialisée
Conversion lysogénique
Conjugaison
Pilus sexuel /F
Réplicon
F+
F-
Hfr
F’
Elément génétique transposable
Séquence d’insertion
Transposon
Recombinaison site-spécifique
Variation de phase
Plasmide, plasmide conjugatif
Plasmide non-conjugatif
Facteur R
RTF
Déterminant R
|
MECHANISME DE TRANSFERT DE GENE CHEZ LES BACTERIES
A. Transformation
La transformation est le transfert de gènes résultant de l’absorption
d’ADN nu par une cellule receveuse à partir d’une cellule donneuse.
Certaines bactéries (ex : Bacillus, Haemophilus, Neisseria, Pneumococcus)
peuvent absorber de l’ADN à partir de l’environnement et l’ADN absorbé peut
être incorporé dans le chromosome du receveur.
1. Facteurs affectant la transformation
a. Taille de l’ADN
L’ADN double brin qui fonctionne le mieux pour la transformation
fait au moins 5 X 105 daltons. En conséquence, la transformation est
sensible aux nucléases de l’environnement.
b. Compétence du receveur
Certaines bactéries sont capables d’absorber de l’ADN naturellement.
Cependant, ces bactéries prennent seulement de l’ADN à un moment
précis de leur cycle de croissance quand elles produisent une
protéine spécifique appelée facteur de compétence. A cette étape on
dit que les bactéries sont compétentes. D’autres bactéries ne sont
pas capables d’absorber de l’ADN naturellement. Cependant, dans ces
bactéries, la compétence peut être induite in vitro par traitement
chimique (ex : CaCl2).
2. Etapes de la transformation
a. Absorption de l’ADN
L’absorption d’ADN diffère pour les bactéries en fonction qu’elles
soient Gram + ou Gram -. Dans les bactéries Gram+ l’ADN est absorbé
sous la forme d’une molécule simple brin et le brin complémentaire
est fabriqué chez le receveur. Par contraste, les bactéries Gram –
absorbent l’ADN sous forme de double brin.
b. Recombinaison légitime/homologue/générale
Après que l’ADN du donneur ait été internalisé, un évènement de
recombinaison réciproque a lieu entre le chromosome et l’ADN donneur.
Cette recombinaison nécessite une homologie entre l’ADN donneur et
le chromosome ce qui résulte en la substitution de l’ADN entre le
receveur et le donneur comme illustré sur la figure 2.
|
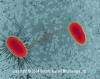 Souches d’E. Coli (bacille procaryote) subissant la
conjugaison. L’une des souche présente un fimbriae.
©
Dr Dennis
Kunkel, University of Hawaii. Used with permission
Souches d’E. Coli (bacille procaryote) subissant la
conjugaison. L’une des souche présente un fimbriae.
©
Dr Dennis
Kunkel, University of Hawaii. Used with permission
 Figure 1 : Gènes de transfert ayant été montrés comme
étant présents dans différentes espèces de bactéries
Figure 1 : Gènes de transfert ayant été montrés comme
étant présents dans différentes espèces de bactéries
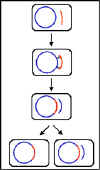 Figure 2 : Recombinaison générale. L’ADN donneur est
représenté en rouge et l’ADN receveur en bleu.
Figure 2 : Recombinaison générale. L’ADN donneur est
représenté en rouge et l’ADN receveur en bleu.
 Figure 3 : Mécanisme de la transduction généralisée
Figure 3 : Mécanisme de la transduction généralisée |
La recombinaison nécessite les gènes de recombinaison
bactériens (recA, B et C) et une homologie entre les ADN
impliqués. Ce type de recombinaison est appelé légitime ou
homologue ou générale. A cause de la nécessité d’une homologie
entre les ADN du donneur et de l’hôte, c’est seulement l’ADN de
bactéries fortement apparentées qui peut se transformer de
manière efficace, bien que dans de rares cas le transfert de
gènes entre des bactéries peu apparentées a déjà été mis en
évidence.
3. Signification
La transformation a lieu dans la nature et peut mener à une
augmentation de la virulence. De plus, la transformation est
largement utilisée dans la technologie de l’ADN recombinant.
B. Transduction
La transduction est le transfert d’information génétique à partir
d’un donneur vers un receveur via un bactériophage. La paroi du phage
protège l’ADN de l’environnement et de cette manière la transduction,
contrairement à la transformation, n’est pas affectée par les nucléases
de l’environnement. Tous les phages ne peuvent pas médier la
transduction. Dans la plupart des cas le transfert de gène a lieu entre
des membres de la même famille de bactéries. Cependant, si un phage
particulier possède une large gamme d’hôtes alors le transfert entre
espèces peut avoir lieu. La capacité d’un phage à médier la transduction
est reliée au cycle de vie du phage.
1. Types de transduction
a. Transduction généralisée
La transduction généralisée est une transduction au cours de
laquelle potentiellement n’importe quel gène bactérien de
donneur peut être transféré à un receveur. Le mécanisme de la
transduction généralisée est illustré dans la figure 3.
Les phages qui permettent la transduction généralisée découpent
généralement l’ADN de l’hôte en plus petits morceaux et
encapsident cet ADN dans une particule phagique par un mécanisme
d’encapsidation fortuite. Occasionnellement, un des morceaux de
l’ADN de l’hôte est accidentellement empaqueté dans la capside
du phage. Ainsi, n’importe quel gène du donneur peut
potentiellement être transféré mais ne sera transféré qu’autant
d’ADN que peut contenir une tête de phage. Si une cellule
réceptrice est infectée par un phage qui contient de l’ADN du
donneur, l’ADN du donneur entre dans le receveur. Dans la
cellule réceptrice un évènement de recombinaison généralisée qui
substitue l’ADN du donneur à l’ADN du receveur peut avoir lieu (voir
figure 2).
|
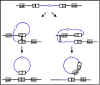 Figure 4 : Mécanisme de la transduction spécifique
Figure 4 : Mécanisme de la transduction spécifique |
b. Transduction spécialisée
La transduction spécialisée est une transduction au cours de
laquelle seulement certains gènes du donneur peuvent être
transférés au receveur. Différents phages peuvent transférer
différents gènes mais un phage individuel peut seulement
transférer certains gènes. La transduction spécialisée est
médiée par des phages lysogéniques ou tempérés et les gènes
qui sont transférés dépendent d’où le prophage est inséré
dans le chromosome. Le mécanisme de la transduction
spécialisée est illustré dans la figure 4.
Pendant
l’excision du prophage, occasionnellement une erreur peut se
produire quand une partie de l’ADN de l’hôte est excisée
avec celui du phage. C’est seulement l’ADN de l’hôte situé
de part et d’autre de l’endroit où le phage est inséré qui
peut être transféré (d’où la transduction spécialisée).
Après réplication et libération du phage et infection du
receveur, la lysogénisation du receveur peut avoir lieu, ce
qui résulte en un transfert stable des gènes du donneur. Le
receveur va alors avoir deux copies des gènes qui ont été
transférés. La recombinaison légitime de gènes entre le
donneur et le receveur est aussi possible.
2. Signification
La conversion lysogénique (phagique) a lieu dans la nature et
est la source de souches virulentes de bactéries.
|
|
MOVIE
Conjugation
High
resolution
Low resolution
© Mondo Media, San Francisco, Calif., USA and
and The
MicrobeLibrary
This video clip demonstrates the process of conjugation. First, two bacteria combine via a sex pilus. Next, one strand of the plasmid is transferred to the attached cell. Note that the original plasmid is not lost from the first cell. Finally, each cell immediately duplicates the single strand so that both bacteria have
a copy of the double stranded plasmid
|
C. Conjugaison
Il s’agit du transfert d’ADN d’un donneur à un receveur par contact physique
direct entre les cellules. Chez les bactéries il y a deux types de
reproducteurs, un donneur (mâle) et un receveur (femelle) et la direction de
transfert du matériel génétique est unidirectionnelle ; l’ADN est transféré
du donneur au receveur.
1. Types de reproducteurs chez
les bactéries
a. Donneur
La capacité d’une bactérie à être un donneur est la conséquence de
la présence dans la cellule d’un morceau supplémentaire d’ADN appelé
facteur F ou facteur de fertilité ou facteur sexuel. Le facteur F
est un morceau d’ADN circulaire qui peut se répliquer de manière
autonome dans la cellule ; c’est un réplicon indépendant. Les
morceaux d’ADN extrachromosomiques qui peuvent se répliquer de
manière autonome ont le nom générique de plasmide. Le facteur F
porte des gènes qui sont nécessaires à sa réplication et pour sa
capacité à transférer de l’ADN au receveur. L’une des choses pour
laquelle code le facteur F est la possibilité de produire un pilus
sexuel (pilus F) à la surface de la bactérie. Ce pilus est important
pour le processus de conjugaison. Le facteur F n’est pas seulement
un plasmide qui peut médier la conjugaison mais il est généralement
utilisé comme modèle.
b. Receveur
La capacité à agir comme un receveur est une conséquence de
l’absence du facteur F.
|
| |
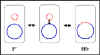 a
a
 b
b
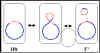
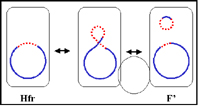
Figure 5 : Etats physiologiques du facteur F.
|
2. Etats physiologiques du facteur F
a. Autonome (F+)
Dans cet état le facteur F porte seulement les gènes
nécessaires pour sa réplication et pour le transfert d’ADN.
Il n’y a pas de gène chromosomique associé avec le facteur F
dans les souches F+.
Lors de croisements entre les types F+ et F- le F-
devient F+ alors que le F+ reste F+. Ainsi, le facteur F est
infectieux. De plus, il y a seulement un faible taux de
transfert de gènes chromosomiques.
b. Intégré (Hfr)
Dans cet état le facteur F est intégré dans le chromosome
bactérien via un évènement de recombinaison comme illustré
sur la figure 5a.
Lors de croissements entre les types Hfr et F- le F- devient
rarement Hfr et Hfr reste Hfr. De plus, il existe une forte
fréquence de transfert de gènes chromosomiques du donneur.
|
| |
c. Autonome avec gènes chromosomiques (F’)
Dans cet état le facteur F est autonome mais il porte cette
fois quelques gènes chromosomiques. Les facteurs F’ sont
produits par excision du facteur F d’un Hfr, comme illustré
sur la figure 5b. Occasionnellement quand le facteur F est
excisé d’un chromosome Hfr, les gènes du donneur situés de
part et d’autre du facteur F peuvent être excisés avec le
facteur F générant un F’. Les facteurs F’ sont nommés selon
les gènes chromosomiques qu’ils portent.
Lors de
croisements entre F’ et F- le F- devient F’ alors que le F’
reste F’. De plus il y a de fortes fréquences de transfert
des gènes chromosomiques de F’ et de faibles fréquences de
transfert des autres gènes chromosomiques du donneur.
|
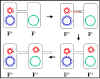 Figure 6 : Mécanisme de croisement F+ x F-
Figure 6 : Mécanisme de croisement F+ x F- |
3. Mécanisme de conjugaison
a. Croisement F+ x F-
i) Formation de pair
Le bout du pilus sexuel vient au contact du receveur et
un pont de conjugaison se forme entre les deux cellules.
C’est par ce pont que l’ADN va passer du donneur vers le
receveur. Ainsi, l’ADN est protégé des nucléases de
l’environnement. Les couples appariés peuvent être
séparés par des forces de cisaillement et la conjugaison
est alors interrompue. Ainsi les couples appariés
restent associées seulement pour un court moment.
ii) Transfert d’ADN
L’ADN plasmidique est entaillé à un site spécifique
appelé origine de transfert et est répliqué par un
mécanisme de « cercle roulant ». Un ADN simple brin
passe dans le pont de conjugaison et entre dans le
receveur où le second brin est synthétisé.
iii) Ce processus explique les caractéristiques du
croisement F+ x F-. Le receveur devient F+, le donneur
reste F+ et il y a un faible taux de transfert de gènes
chromosomiques du donneur. En effet, comme décrit dans
la figure 7 il n’y a pas de transfert de gènes
chromosomiques du donneur. Dans la pratique cependant,
il y a un faible taux de transfert de gènes
chromosomiques du donneur lors de ces croisements.
|
|
ANIMATION
Mating of F+ and F- Bacterial Strains
© Thomas M. Terry, University of Connecticut, Storrs, Conn., USA
and
The
MicrobeLibrary
The F plasmid is a self-transmissible plasmid found in some strains of E. coli. Cells that possess one or more copies of the F plasmid are called F+; cells lacking the F plasmid are called F-. The animation illustrates several stages in the transfer of the F plasmid from F+ to F- cells.
|
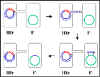 Figure 7 : Mécanisme de croisement Hfr x F-
Figure 7 : Mécanisme de croisement Hfr x F- |
b. Croisement Hfr x F- (Figure 7)
i) Formation de paire
ii) Transfert d’ADN
L’ADN est entaillé au niveau de l’origine de transfert
et est répliqué par un mécanisme de cercle roulant. Mais
l’ADN qui est transféré est le chromosome. En fonction
d’où le facteur F s’est intégré dans le chromosome et de
son orientation, différents gènes chromosomiques seront
transférés à différents moments. Cependant, l’ordre et
les distances relatives des gènes resteront toujours les
mêmes. C’est seulement quand l’ensemble du chromosome
est transféré que le facteur F est transféré. Puisque
des forces de cisaillement séparent les couples appariés
il est rare que l’ensemble di chromosome soit transféré.
Ainsi, le receveur ne reçoit pas le facteur F lors d’un
croisement Hfr x F-.
iii) Recombinaison légitime
La recombinaison entre l’ADN transféré et le chromosome
résulte en l’échange de matériel génétique entre le
donneur et le receveur.
iv) Ce mécanisme explique les caractéristiques du
croisement Hfr x F-. Le receveur reste F-, le donneur
reste Hfr et il y a une forte fréquence de transfert de
gènes chromosomiques du donneur.
|
|
ANIMATION
Mating of Hfr and F- Bacterial Strains
© Thomas M. Terry, University of Connecticut, Storrs, Conn., USA
and
The
MicrobeLibrary
|
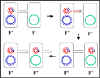 Figure 8 : Mécanisme de croisement F’ x F-
Figure 8 : Mécanisme de croisement F’ x F- |
c. Croisement F’ x F- (Figure 8)
i) Formation de paire
ii) Transfert d’ADN
Ce processus est similaire au croisement F+ x F-. Cependant, comme
le F’ porte quelques gènes chromosomiques ils seront aussi
transférés.
iii) La recombinaison homologue n’est pas nécessaire
bien qu’elle puisse se produire.
iv) Ce mécanisme explique les caractéristiques du
croisement F’ x F-. Le F- devient F’, le F’ reste F’ et il y a une
forte fréquence de transfert de gènes du donneur sur le F’ mais une
faible fréquence de transfert des autres gènes chromosomiques du
donneur.
4. Signification
Parmi les bactéries Gram négatives il y a une voie
majeure selon laquelle les gènes sont transférés. Le transfert peut se
produire entre différentes espèces de bactéries. Le transfert de
résistances multiples aux antibiotiques par conjugaison est devenu un
problème majeur dans le traitement de certaines maladies bactériennes.
Puisque la cellule receveuse devient un donneur après transfert d’un
plasmide il est aisé de voir pourquoi un gène de résistance aux
antibiotiques porté sur un plasmide peut rapidement convertir une
population de cellules sensibles en une population résistante.
Les bactéries à Gram positif possèdent aussi des
plasmides qui portent des gènes de résistances multiples aux
antibiotiques, dans certains cas ces plasmides sont transférés par
conjugaison alors que d’autres sont transférés par transduction. Le
mécanisme de conjugaison chez les bactéries à Gram positif est différent
de celui des bactéries à Gram négatif. Chez les bactéries à Gram positif,
le donneur fabrique un matériau adhésif qui cause l’agrégation avec le
receveur et l’ADN est transféré.
|
| |
ELEMENTS GENETIQUES TRANSPOSABLES
A. Eléments
génétiques transposables
Les éléments génétiques transposables sont des segments
d’ADN qui ont la capacité de bouger à partir d’une position à une autre.
B. Propriétés des éléments
génétiques transposables
1. Mouvement aléatoire
Les éléments génétiques transposables peuvent bouger
de n’importe quelle molécule d’ADN à une autre molécule d’ADN ou
même à un autre endroit de la même molécule. Le mouvement n’est pas
totalement aléatoire ; il y a des sites de préférence dans la
molécule d’ADN auxquels les éléments génétiques transposables vont
s’insérer.
2. Incapable
d’autoréplication
Les éléments génétiques transposables n’existent pas
de manière autonome (exception – quelques phages transposables) et
ainsi, pour être répliqués ils doivent faire partie d’un autre
réplicon.
3. Transposition médiée par la
recombinaison site-spécifique
La transposition nécessite peu ou pas d’homologie
entre sa position de départ et le nouveau site. L’évènement de
transposition est médié par une transposase codée par l’élément
génétique transposable. La recombinaison qui ne nécessite pas
d’homologie entre les molécules se recombinant est appelée site-spécifique
ou illégitime ou homologue.
4. La transposition peut être accompagnée
d’une duplication
Dans de nombreux cas la transposition d’un élément
génétique transposable résulte en le retrait de l’élément de son
site d’origine et en l’insertion dans un nouveau site. Cependant,
dans certains l’évènement de transposition est accompagné d’une
duplication de l’élément génétique transposable. Une copie reste au
site d’origine et l’autre est transposée au nouveau site.
|
 Figure 9 : Structure d’éléments génétiques transposables
Figure 9 : Structure d’éléments génétiques transposables |
C. Types d’éléments génétiques
transposables
1. Séquences d’insertion (IS)
Les séquences d’insertion sont des éléments génétiques transposables
qui ne portent pas de gène connu sauf ceux nécessaires à la
transposition.
a. Nomenclature
Les séquences d’insertion sont appelées IS suivi d’un numéro ; ex :
IS1
b. Structure (figure 9)
Les séquences d’insertion sont de courts brins d’ADN qui possèdent à
leurs extrémités des séquences répétées, qui sont impliquées dans la
transposition. Entre les séquences répétées terminales se trouvent
des gènes impliqués dans la transposition et des séquences qui
contrôlent l’expression de ces gènes mais aucun autre gène non
essentiel n’est présent.
c. Importance
i) Mutation
L’introduction d’une séquence d’insertion dans un gène bactérien
résulte en l’inactivation de ce gène.
ii) Insertion de plasmide dans les chromosomes
Les sites auxquels s’insèrent les plasmides dans le chromosome
bactérien sont au niveau ou près d’une séquence d’insertion dans
le chromosome.
iii) Variation de phase
Les antigènes du flagelle sont les principaux antigènes contre
lesquels le système immunitaire est dirigé dans le but de
combattre l’infection bactérienne. Chez Salmonelle il existe
deux gènes qui codent pour deux antigènes du flagelle
antigéniquement différents. L’expression de ces gènes est
régulée par une séquence d’insertion. Dans une orientation un
des gènes est actif alors que dans l’autre orientation l’autre
gène du flagelle est actif. Ainsi, Salmonella peut changer son
flagelle en réponse aux attaques du système immunitaire. La
variation de phase n’est pas unique aux antigènes de flagelle de
Salmonella. Elle est également observée pour d’autres antigènes
de surface de bactéries. Cependant le mécanisme de variation de
phase puisse différer dans différentes espèces de bactéries (ex
: Neisseria ; transformation).
2. Transposons (Tn)
Les transposons sont des éléments génétiques transposables qui
portent un ou plus d’autres gènes en plus de ceux qui sont essentiels à
la transposition.
|
 Figure 10 : Structure d’un transposon
Figure 10 : Structure d’un transposon |
a. Nomenclature
Les transposons sont désignés par Tn suivit d’un numéro.
b. Structure
La structure d’un transposon est similaire à une séquence
d’insertion. Les gènes supplémentaires sont situés entre les
séquences terminales répétées. Dans certains cas (transposons
composites) les séquences terminales répétées sont en fait
des séquences d’insertion. (Voir figure 10).
c. Importance
De nombreux gènes de résistance aux antibiotiques sont
situés sur des transposons. Puisque les transposons peuvent
sauter d’une molécule d’ADN à une autre, ces transposons
portant des résistances aux antibiotiques sont un facteur
majeur du développement de plasmides qui peuvent conférer
une résistance multiple à la bactérie qui porte un tel
plasmide. Ces plasmides de résistance multiple sont devenus
un problème médical majeur à cause de l’utilisation à tort
et à travers des antibiotiques et ont conféré un avantage
sélectif pour les bactéries portant ces plasmides.
|
| |
PLASMIDES
A. Définition
Les plasmides sont des éléments génétiques extrachromosomiques capables
d’une réplication autonome. Un épisome est un plasmide qui peut s’intégrer
dans un chromosome bactérien.
B. Classification des plasmides
1. Propriétés de transfert
a. Plasmides conjugatifs
Les plasmides conjugatifs sont ceux qui permettent la conjugaison.
Ces plasmides sont généralement grands et ont tous les gènes
nécessaires pour la réplication autonome et pour le transfert d’ADN
au receveur (ex : gènes du pilus sexuel).
b. Plasmides non-conjugatifs
Les plasmides non–conjugatifs sont ceux qui ne peuvent pas médier la
conjugaison. Ils sont généralement plus petits que les plasmides
conjugatifs et il leur manque un ou plusieurs gènes nécessaires pour
le transfert d’ADN. Un plasmide non-conjugatif peut être transféré
par conjugaison si la cellule porte aussi un plasmide conjugatif.
2. Effets phénotypiques
a. Plasmide de fertilité (Facteur F)
b. Plasmides bactériocinogéniques
Ces plasmides portent des gènes qui codent pour une substance qui
tue les autres bactéries. Ces substances sont appelées bactériocines
ou colicines.
c. Plasmides de résistance (facteurs R)
Ces plasmides portent des gènes de résistance aux antibiotiques.
i) Origine - L’origine des facteurs R n’est pas connue. Il
est probable qu’ils ont évolué pour d’autres raisons et que
l’avènement de l’ère des antibiotiques a fourni un avantage
sélectif pour leur large dissémination.
|
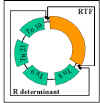 Figure 11 : Structure d’un plasmide R
Figure 11 : Structure d’un plasmide R |
ii) Structure - Les plasmides R sont des plasmides
conjugatifs dans lesquels les gènes pour la réplication et
le transfert sont localisés sur une partie du facteur R et
les gènes de résistance sont localisés sur une autre partie
comme illustré dans la figure 11.
RTF (Facteur de Résistance Transfert)- porte les gènes de
transfert.
Déterminant R- porte les gènes de résistance. Les gènes de
résistance sont souvent des parties de transposons.
Mode d’action des gènes de résistance:
-
Modification (détoxification) des
antibiotiques – ex : β-lactamase.
-
Altération de la cible – ex : Résistance
à la streptomycine.
-
Altération de l’absorption – résistance
à la tétracycline.
-
Remplacement de la voie sensible – ex :
nouvelle voie de l’acide folique pour la résistance aux
drogues sulfamides.
|
|
|
 Retourner à la section d'bacteriologie de Microbiologie et Immunologie On-line
Retourner à la section d'bacteriologie de Microbiologie et Immunologie On-line
This page last changed on
Thursday, September 05, 2013
Page maintained by
Richard Hunt
Please report any problems to
richard.hunt@uscmed.sc.edu
|
 Souches d’E. Coli (bacille procaryote) subissant la
conjugaison. L’une des souche présente un fimbriae.
©
Dr Dennis
Kunkel, University of Hawaii. Used with permission
Souches d’E. Coli (bacille procaryote) subissant la
conjugaison. L’une des souche présente un fimbriae.
©
Dr Dennis
Kunkel, University of Hawaii. Used with permission
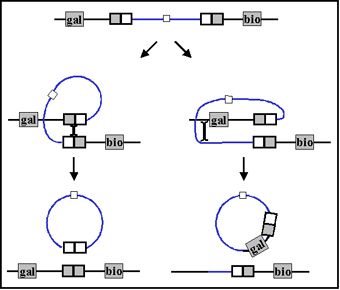 Figure 4 : Mécanisme de la transduction spécifique
Figure 4 : Mécanisme de la transduction spécifique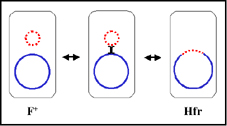 a
a
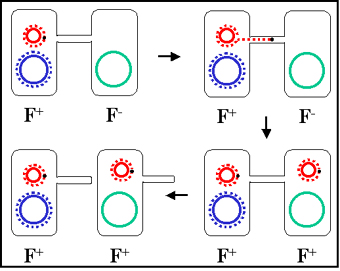 Figure 6 : Mécanisme de croisement F+ x F-
Figure 6 : Mécanisme de croisement F+ x F-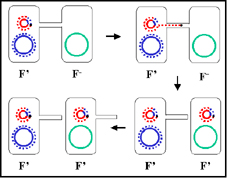 Figure 8 : Mécanisme de croisement F’ x F-
Figure 8 : Mécanisme de croisement F’ x F-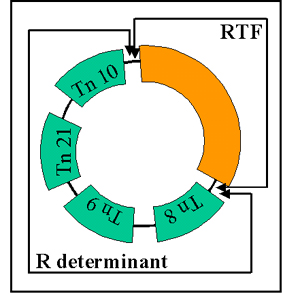 Figure 11 : Structure d’un plasmide R
Figure 11 : Structure d’un plasmide R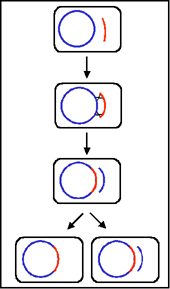
{kind=link}
{kind=link}