| x | x | |||||||||
|
|
 |
|||||||||
|
|
BACTERIOLOGÍA | INMUNOLOGÍA | MICOLOGÍA | PARASITOLOGÍA | VIROLOGÍA | |||||
|
|
||||||||||
|
VIDEOCONFERENCIA EN INGLÉS |
||||||||||
| CENTRO DE INVESTIGACION Y DE ESTUDIOS AVANZADOS DEL IPN | ||||||||||
| BUSCAR | ||||||||||
|
Let us know what you think FEEDBACK |
||||||||||
|
Editado e ilustrado por Dr Richard Hunt |
||||||||||
|
OBJETIVOS DE ENSEÑANZA Describir la inmunidad específica y las células que participan
|
DESCRIPCIÓN GENERAL
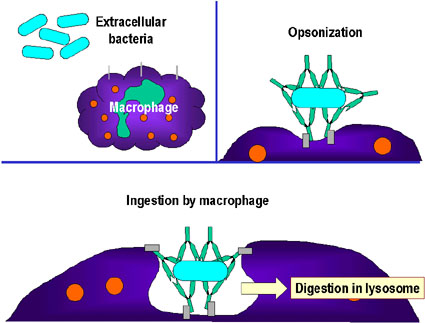
Deberá notarse que cada clase de anticuerpo tiene distintos sitios de acción y no son igualmente efectivos en neutralización, opsonización o en la activación del complemento. Los anticuerpos no son particularmente efectivos contra patógenos que residen intracelularmente.
|
|||||||||
|
Figura 1 |
A
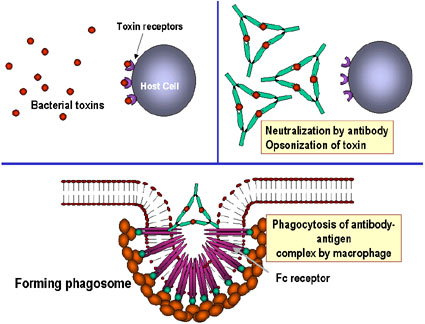
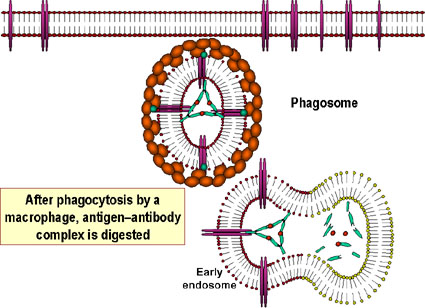
Unión y neutralización de una toxina bacteriana por anticuerpos, impidiendo la interacción con el hospedero y el desarrollo de la patología. La toxina libre puede reaccionar con los receptores de la célula hospedera, no así los complejos toxina-anticuerpo. Los anticuerpos también neutralizan completamente partículas virales y células bacterianas al unírseles a estas e inactivándolas. El complejo antígeno:anticuerpo es eventualmente retirado de la circulación y degradado por los macrófagos. Al recubrir los anticuerpos al antígeno, lo hacen reconocible como extraño por los fagocitos (macrófagos y leucocitos polimorfonucleares), los cuales lo ingieren y lo destruyen; este proceso se denomina opsonización. B
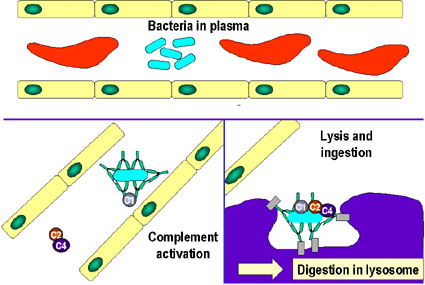 Activación
del sistema del complemento por anticuerpos que recubren una célula
bacteriana. Los anticuerpos unidos forman un receptor para la primera
proteína del sistema del complemento, la cual eventualmente forma un
complejo proteico en la superficie de las bacterias que en algunos
casos, puede matar directamente a esta pero más generalmente favorece su
eliminación y degradación por fagocitos.
Activación
del sistema del complemento por anticuerpos que recubren una célula
bacteriana. Los anticuerpos unidos forman un receptor para la primera
proteína del sistema del complemento, la cual eventualmente forma un
complejo proteico en la superficie de las bacterias que en algunos
casos, puede matar directamente a esta pero más generalmente favorece su
eliminación y degradación por fagocitos.
|
|||||||||
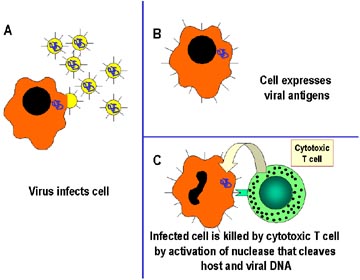 Figura 2 Mecanismos de defensa del
hospedero en contra de una infección intracelular por Virus. Las células
son reconocidas por células T especializadas llamadas Tc o Linfocitos T
citotóxicos (CTLs), los cuales matan directamente a las células
infectadas. El mecanismo de citotoxicidad implica la activación de
nucleasas en la células infectada, las cuales rompen tanto el DNA de la
célula infectada como el viral.
Figura 2 Mecanismos de defensa del
hospedero en contra de una infección intracelular por Virus. Las células
son reconocidas por células T especializadas llamadas Tc o Linfocitos T
citotóxicos (CTLs), los cuales matan directamente a las células
infectadas. El mecanismo de citotoxicidad implica la activación de
nucleasas en la células infectada, las cuales rompen tanto el DNA de la
célula infectada como el viral. |
Respuestas mediadas por células
|
|||||||||
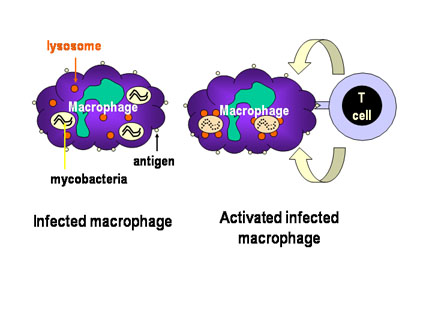 Figura 3
Figura 3Mecanismo de defensa del hospedero contra la infección intracelular por micobacterias. Las micobacterias infectan macrófagos y estas se alojan en las vesículas citoplasmáticas que resisten la fusión con lisosomas y la posterior destrucción de la bacteria por la actividad bactericida del macrófago. Sin embargo cuando la célula Th1 apropiada reconoce un macrófago infectado, esta libera moléculas activadoras de macrófagos que inducen la fusión lisosomal y la activación de la actividad bactericida del macrófago. |
Las células que alojan a los patógenos en el sistema vesicular son reconocidas por una subpoblación de células T cooperadoras, las células Th1 o células T inflamatorias que liberan moléculas llamadas citocinas que capacitan a las células infectadas para matar al patógeno intracellular.
|
|||||||||
|
Poblaciones de células T Las células T participan en los dos tipos de respuestas inmunológicas: la humoral (anticuerpos) y la cellular. Existen subpoblaciones de células T que tienen las siguientes funciones:
|
||||||||||
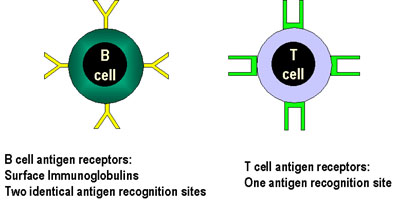 Figura 4
Figura 4El receptor para antígeno de la célula B tiene dos sitios de reconocimiento mientras que el de las células T solo uno. |
Especificidad de la respuesta inmune La especificidad de estas respuestas reside en el receptor de células T (TCR) el cual reconoce péptidos derivados de patógenos que se encuentran unidos al complejo principal de histocompatibilidad (MHC) siendo estas moléculas expresadas en la superficie de células nucleadas. Todos los TCR de una célula individual tienen especificidad única. (RECORDAR: el receptor de la célula B que une antígenos es una inmunoglobulina unida a la membrana, y todas las inmunoglobulinas de una célula B tienen especificidad única.
|
|||||||||
|
Diversidad de la especificidad de los receptores
|
||||||||||
|
Clases de moléculas del Complejo Principal de Histocompatibilidad (MHC) reconocidas por el Receptor de Células T (TCR). El TCR reconoce péptidos unidos a las moléculas del MHC expresadas en la superficie de las células presentadoras de antígeno. Hay dos clases de moléculas MHC, las MHC clase I y las MHC clase II.
NOTA: La fragmentación de las proteínas y la asociación de los péptidos correspondientes con cada una de las dos clases de moléculas MHC efectuada por diferentes vías es lo que se conoce colectivamente como procesamiento y presentación de antígeno.
|
||||||||||
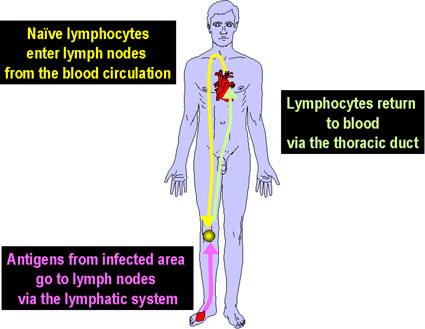 Figura 5
Figura 5Los linfocitos circulantes se encuentran con el antígeno en los tejidos linfoides periféricos |
Recirculación linfocítica
|
|||||||||
|
IMMUNIDAD: CONTRASTE ENTRE LO NO-ESPECÍFICO Y LO ESPECÍFICO
|
||||||||||
| Figura 6 |
CELULAS DEL SISTEMA INMUNE Todos los tipos celulares del sistema inmune se originan en la medula ósea.
|
|||||||||
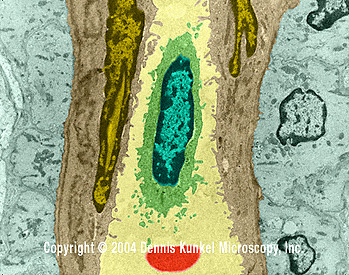
 Linfocito T Humano (SEM x12,080)
©
Dennis
Kunkel Microscopy, Inc.
Used with permission
Linfocito T Humano (SEM x12,080)
©
Dennis
Kunkel Microscopy, Inc.
Used with permission |
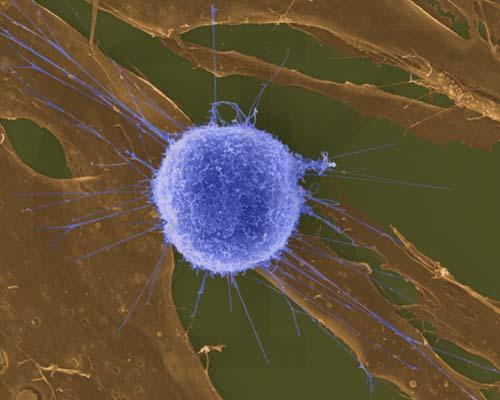 LinfocitoT
Humano Atacando a un Fibroblasto Tumoral/ Célula Cancerosa (SEM x4,000)
©
Dennis Kunkel
Microscopy, Inc.
Used
with permission
LinfocitoT
Humano Atacando a un Fibroblasto Tumoral/ Célula Cancerosa (SEM x4,000)
©
Dennis Kunkel
Microscopy, Inc.
Used
with permission
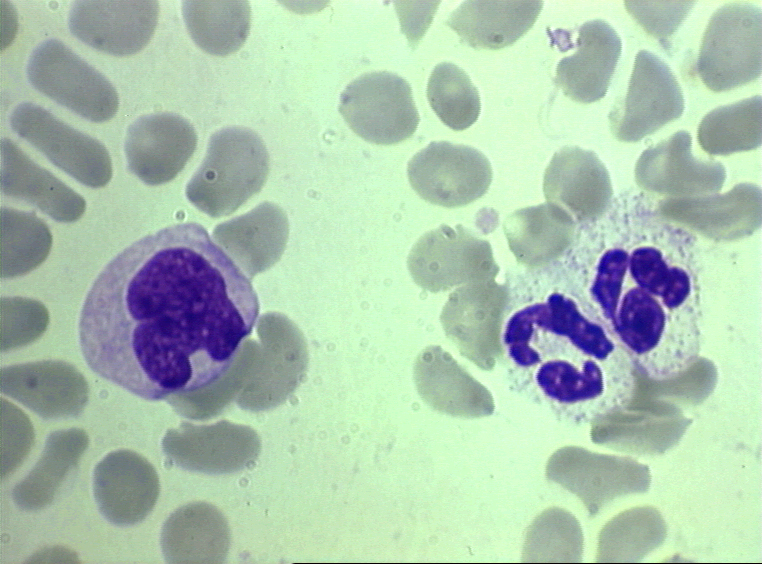
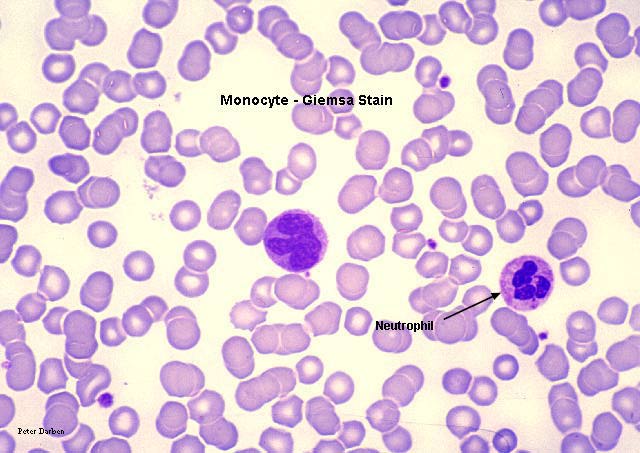 Monocito,
frotis sanguíneo teñido con giemsa
©
Dr
Peter Darben,
Queensland University of Technology clinical parasitology
collection. Used with permission Monocito,
frotis sanguíneo teñido con giemsa
©
Dr
Peter Darben,
Queensland University of Technology clinical parasitology
collection. Used with permission
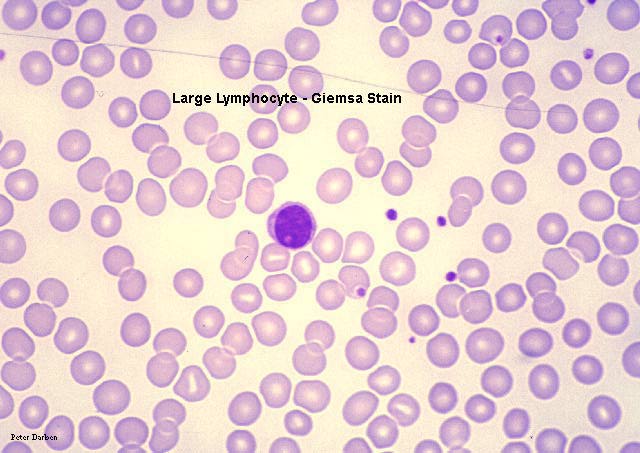
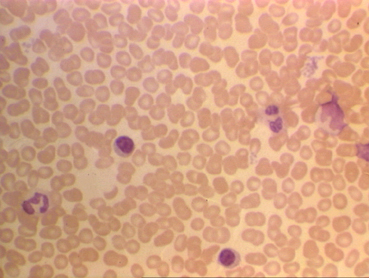 Frotis sanguíneo mostrando linfocitos pequeños
© Bristol Biomedical Image Archive Used with permission
Frotis sanguíneo mostrando linfocitos pequeños
© Bristol Biomedical Image Archive Used with permission
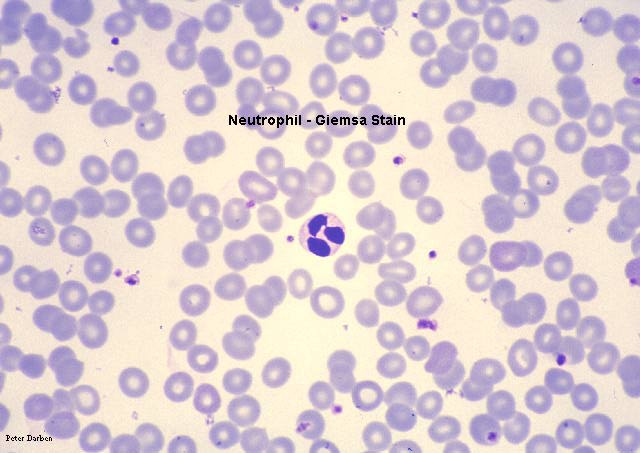
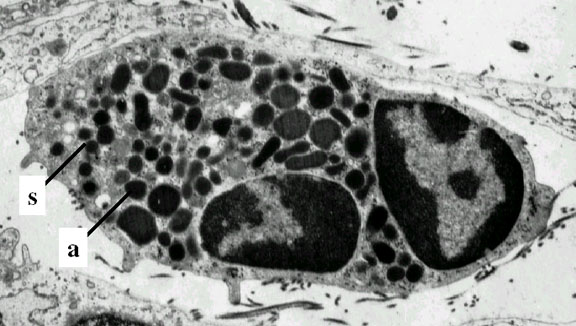 Neutrófilo – micrografía electrónica. Note los dos globules nucleares y
los gránulos azurófilos
© Dr Louise Odor, University of South Carolina School of Medicine
Neutrófilo – micrografía electrónica. Note los dos globules nucleares y
los gránulos azurófilos
© Dr Louise Odor, University of South Carolina School of Medicine
|
|||||||||
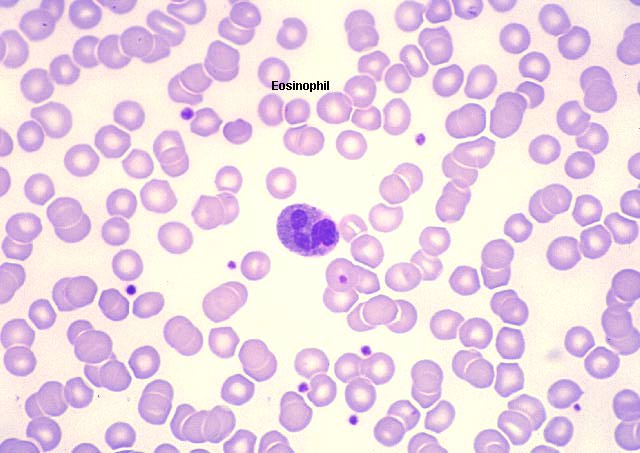
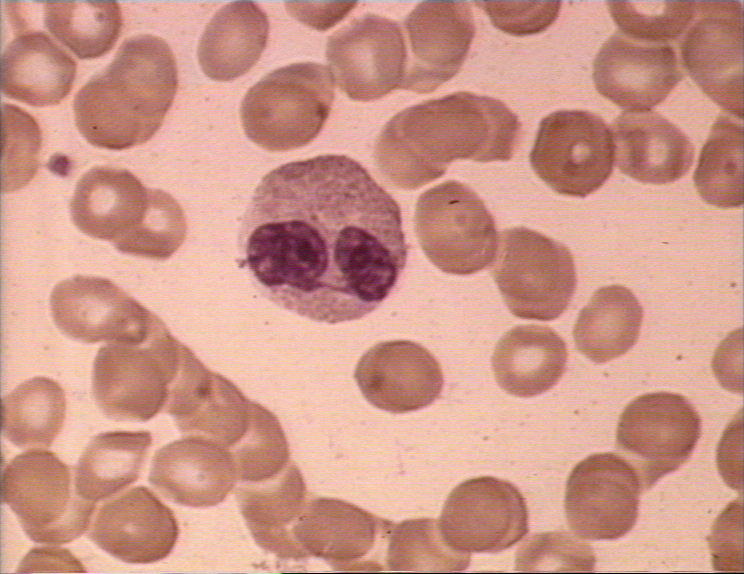 Eosinófilo en frotis sanguíneo
© Bristol Biomedical Image Archive Used with permission
Eosinófilo en frotis sanguíneo
© Bristol Biomedical Image Archive Used with permission
|
||||||||||
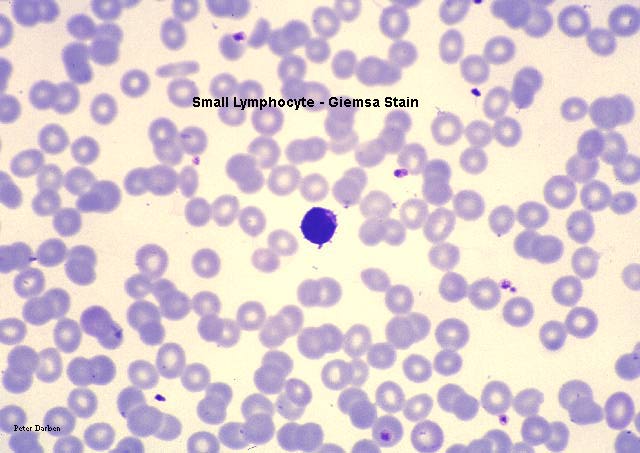 Linfocito pequeño en frotis sanguíneo teñido con giemsa
©
Dr
Peter Darben,
Queensland University of Technology clinical parasitology
collection. Used with permission
Linfocito pequeño en frotis sanguíneo teñido con giemsa
©
Dr
Peter Darben,
Queensland University of Technology clinical parasitology
collection. Used with permission |
||||||||||
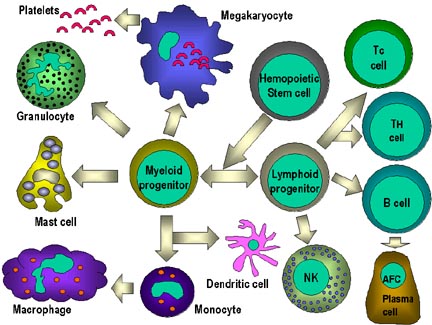
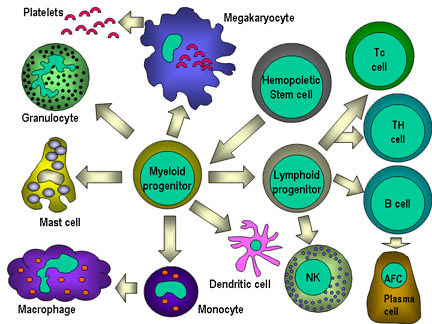 Figura
7
Figura
7Todas las células hematopoyéticas derivan de las células madre pluripotenciales que dan origen a dos principales linajes: uno para células linfoides y otro para células mieloides. El progenitor linfoide común tiene la capacidad de diferenciarse en ya sea células T o células B dependiendo del microambiente en el que se desenvuelven. En mamíferos, las células T se desarrollan en el timo mientras que las células B en el hígado fetal y medula ósea. Una AFC es una célula formadora de anticuerpo, siendo la célula plasmática la AFC más diferenciada. Las células NK también derivan del progenitor linfoide común. Las células mieloides se diferencian en las células comprometidas representadas a la izquierda. El nombre colectivo de granulocitos es usado para eosinófilos, neutrófilos y basófilos |
Hay dos linajes principales que derivan de la célula madre hematopoyética:
MIGRACIÓN LINFÁTICA Y LOCALIZACIÓN Las interacciones celulares productivas que conducen a las respuestas inmunes específicas ocurren principalmente en los ganglios linfáticos y en el bazo. (Los ganglios linfáticos y el bazo son referidos como tejidos linfoides secundarios; la medula ósea y el timo son referidos como tejidos linfoides primarios). |
|||||||||
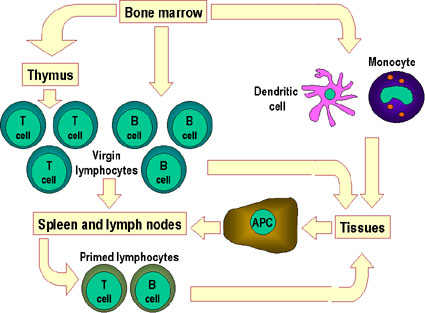 Figura 8.
Los linfocitos vírgenes de los tejidos linfoides primarios tales como la
medula ósea migran hacia los tejidos linfoides secundarios, por ejemplo,
el bazo y los ganglios linfáticos. Las células presentadoras de
antígenos (APCs), que incluyen a las células dendríticas y a los
fagocitos mononucleares los cuales también derivan de las células madre
de la medula ósea. Estas APCs entran en los tejidos capturan al antigeno
y lo transportan hacia los tejidos linfoides para que sean presentados a
las células T y a las células B. los linfocitos activados ahora migran
de los tejidos linfoides y se acumulan predominantemente en los sitios
de infección y de inflamación
Figura 8.
Los linfocitos vírgenes de los tejidos linfoides primarios tales como la
medula ósea migran hacia los tejidos linfoides secundarios, por ejemplo,
el bazo y los ganglios linfáticos. Las células presentadoras de
antígenos (APCs), que incluyen a las células dendríticas y a los
fagocitos mononucleares los cuales también derivan de las células madre
de la medula ósea. Estas APCs entran en los tejidos capturan al antigeno
y lo transportan hacia los tejidos linfoides para que sean presentados a
las células T y a las células B. los linfocitos activados ahora migran
de los tejidos linfoides y se acumulan predominantemente en los sitios
de infección y de inflamación |
La Figura 8 muestra los varios tipos de migración de leucocitos:
|
|||||||||
|
|
|
|||||||||
{kind=link}