![]()

Dr Richard Hunt
VIROLOGÍA
CAPÍTULO DIECIOCHO
LOS VIRUS DE LA HEPATITIS
VIDEOCONFERENCIA
EN INGLÉS
 Figura 1A
Figura 1AVirus de Hepatitis A
CDC
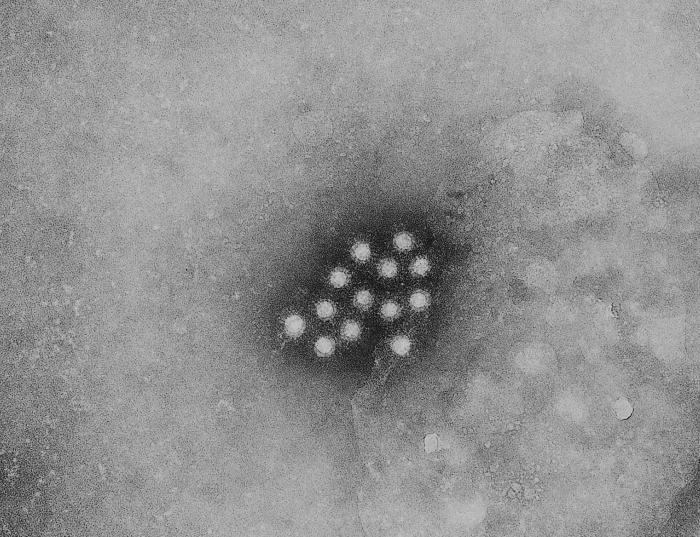 Figura 1B
Figura 1BMicrografía de electrones de un virus de Hepatitis A (VHA)
CDC - Betty Partin
VIRUS DE LA HEPATITIS A
Este picornavirus (figura 1) es el agente causal de la hepatitis infecciosa. Los picornavirus son de cadena sencilla, con una poliadenilación 3’, y un genoma de ARN de sentido positive rodeado por una cápside icosaédrica desnuda (no envuelta) que mide cerca de 28nm de diámetro (figura 2). En el terminal 5’ de la hebra de ARN hay una proteína viral llamada VPg. Solo hay un serotipo para el VHA.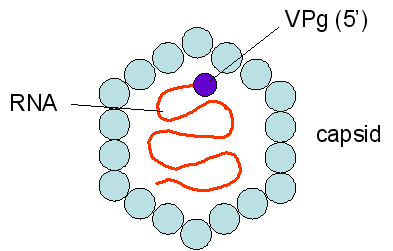 Figura 2
Figura 2Virus de Hepatitis A - un picornavirus
El virus se une a un receptor que se encuentra en la superficie de los hepatocitos y en algunas otras células. El receptor celular 1 del VHA (havcr-1) tiene un ectodominio que contiene una región N-terminal de tipo inmunoglobulina rica en cisterna, continuada por una región de tipo mucina que se extiende desde la primera hasta más allá de la superficie celular. La región de tipo inmunoglobulina se requiere para la unión del VHA. El virus pasa su vida entera en el citoplasma en donde se replica usando una ARN polimerasa ARN dependiente codificada por el mismo virus. Para mayor información sobre la replicación de los picornavirus refiérase a la Sección de Virología Capítulo Cuatro.
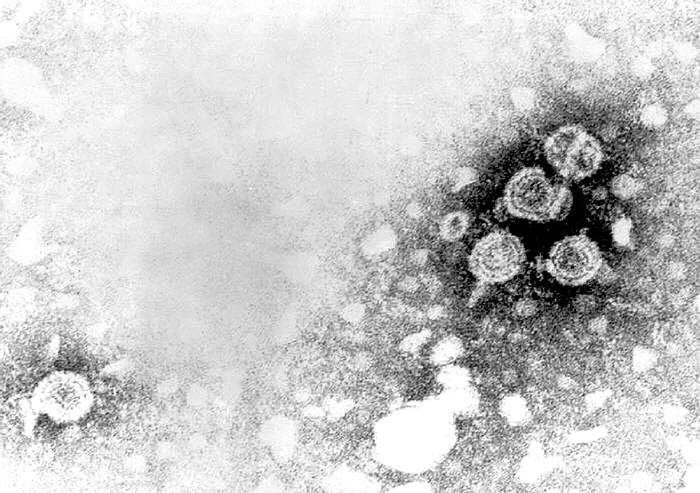 Figura 3A
Figura 3AMicrografía de transmisión de electrones de viriones de hepatitis B, también conocidas como partículas Dane
CDC/Dr. Erski Palmer
![]() Figura 3B
Figura 3B
Virus de Hepatitis B
©
Dra. Linda Stannard,
Universidad de Cape Town, Sudáfrica. Usado con autorización
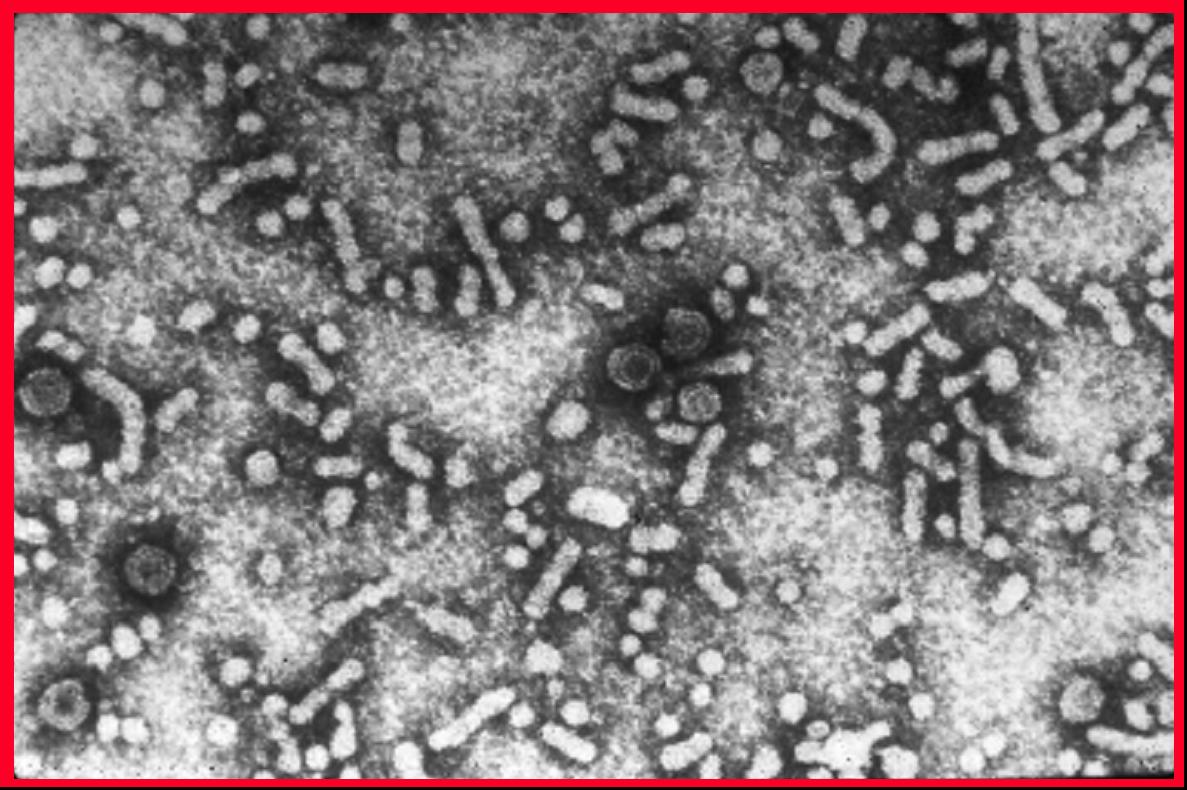 Figura 3C
Figura 3C
Virus de la Hepatitis B
CDC
VIRUS DE LA HEPATITIS B
El virus humano de la Hepatitis B (figura 3) es el prototipo de la familia de los hepadnavirus y causa hepatitis sérica. El VHB tiene un diámetro de aproximadamente 40nm. Infecta a humanos y a chimpancés pero también hay miembros de esta familia de virus que afectan a otros mamíferos y a aves. El VHB es un virus de AND y es envuelto. El ADN es parcialmente de doble cadena y forma un círculo de más o menos 3,200 bases. Aunque está cubierto por una envoltura derivada de la célula huésped, el VHB es extraordinariamente estable a solventes orgánicos. También es resistente a temperatura y a pH. El genoma se asocia con la proteína P (polimerasa) y este complejo es, a su vez, cubierto por antígenos del core (HBcAg y HBeAg). Estas dos proteínas tienen mucho en común en sus secuencias y la mayoría del HBeAg es secretado puesto que es procesado de una forma diferente del HBcAg y por ello no es ensamblado en la progenie del virus. Embebidas en la membrana de bichada lipídica está el antígeno de superficie (HBsAg). El HBsAg (antígeno australiano) está hecho de tres glicoproteínas que son codificadas por el mismo gen Las proteínas se traducen en el mismo marco de lectura y aunque comienzan en un codón AUG diferente; tienen el mimo terminal C. La proteína más grande es la proteína L (42kd) y en su interior contiene la glicoproteína M. La glicoproteína S (27kD) se encuentra dentro de la proteína M. La proteína HBsAg también se secreta al suero del paciente en donde puede ser vista como una partícula esférica (auto asociada con la proteína S) o como partículas filamentosas (también de proteína S, pero también de L y M). Las primeras son más pequeñas que el virus verdadero pero los filamentos pueden ser bastantes largos (varios cientos de nanómetros). Esta gran cantidad de HBsAg libre es responsable de la inhabilidad para detectar anticuerpos contra la proteína al principio de la infección (la llamada “ventana” entre la presencia del HBsAg (indicativo de la presencia del virus) y la presencia de anti-HBsAg).
Las glicoproteínas en la superficie del virus contienen determinantes antigénicos que son grupo-específicos y tipo – específicos. Usando estos determinantes, los epidemiólogos han identificado ocho subtipos del VHB. Los viriones del VHB también se conocen como partículas de Dane.
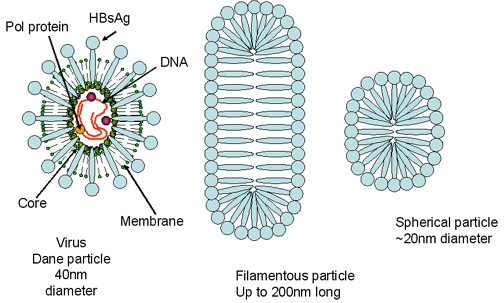 Figura 3D Virus de la Hepatitis B. Partícula de Dane y partículas
incompletes que son halladas en el suero del paciente
Figura 3D Virus de la Hepatitis B. Partícula de Dane y partículas
incompletes que son halladas en el suero del paciente
 Figura 3E
Figura 3E
Estructura del Virus de Hepatitis B
©
Dra. Linda Stannard,
Universidad de Cape Town, Sudáfrica. Usado con autorización
 i
i
 ii
ii
 iii
iii
Figura 4A
Replicación del virus de Hepatitis B
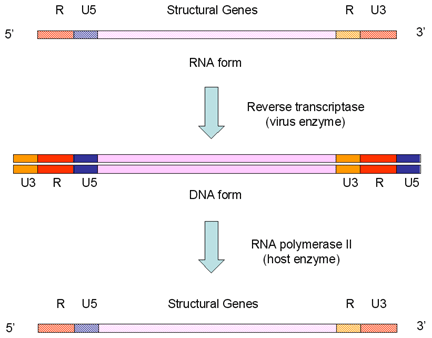 Figura 4B
Figura 4B
Replicación del genoma en los retrovirus
 i
i
 ii
ii
Figura 4C
Replicación del genoma en los hepadnavirus
Replicación
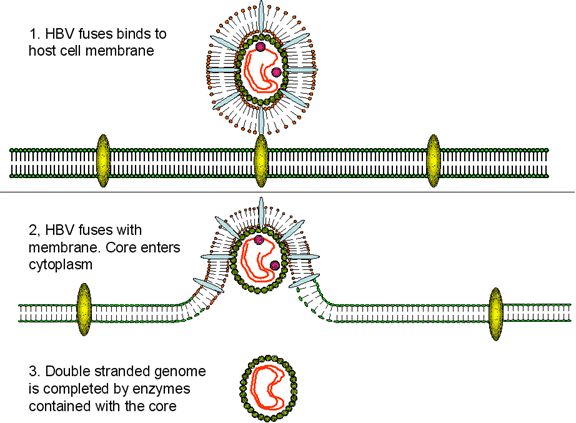
El VHB tiene una forma muy curiosa de replicarse dado que (figura 4A), aún
cuando es un virus de ADN, usa un intermediario de ARN que ha de ser copiado
de vuelta a ADN. El copiado del ARN a ADN no es una función normal de las
células no infectadas pero sí es una función de los retrovirus que también
tienen un genoma de ARN y un intermediario de AND que se integra a los
cromosomas de la célula huésped. Para fines del copiado de ARN a AND, los
retrovirus y el VHB tienen una ADN polimerasa (P) codificada por los mismos
virus llamada transcriptasa inversa.
Luego de que el VHB se fija a s receptor en la superficie celular (el cual aún no ha sido identificado pero podría ser miembro de la familia ovalbúmina de inhibidores de la serina proteasa), la membrana viral se funde con la membrana celular liberando el núcleo (core) al citoplasma. Las proteínas del núcleo (core) se disocian del ADN de cadena parcialmente doble. La ADN polimerasa termina el ADN haciéndolo completamente de cadena doble. Esto se lleva a cabo mediante una polimerasa, codificada por el virus en el citoplasma, que es una de las proteínas del core (a diferencia de la ADN polimerasa celular la cual se encuentra en el núcleo). El ADN de cadena doble entra al núcleo y sus terminales son ligados por enzimas del huésped de manera tal que el virus se vuelve un episoma circular. El ADN viral se asocia con histonas del huésped y se transcribe a ARNm mediante una ARN polimerasa II del huésped. A diferencia de la situación encontrada con los retrovirus, no obstante, la forma de ADN del VHB usualmente no es integrada en el ADN circular; más bien se encuentra como un episoma independiente. Esto es porque, contrario a los retrovirus, los hepadnavirus no tienen actividad integrasa. Sin embargo, las partes integradas del genoma del VHB se encuentran en los cromosomas de muchos pacientes con carcinoma hepatocelular.
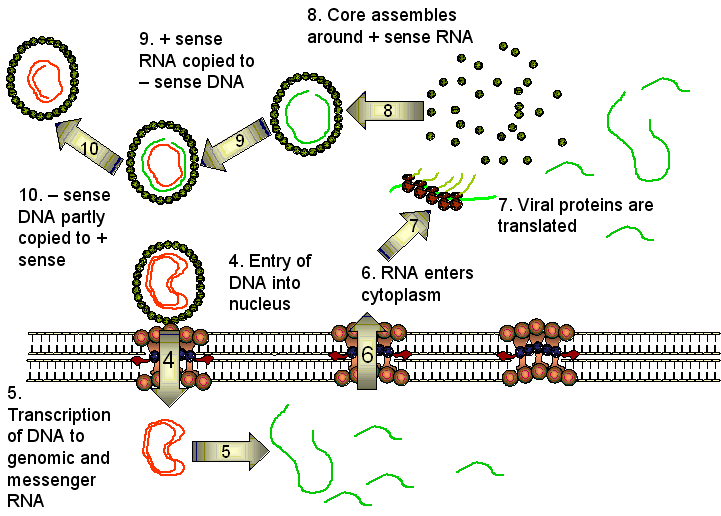
Se sintetizan cuatro ARNm del genoma del VHB. La ARN polimerasa del huésped interactúa con cuatro promotores pero la transcripción siempre cesa en el mismo sitio de poliadenilación de modo tal que los ARN que solapan tienen un terminal 3’ en común. Uno de estos ARNm es ligeramente más largo que la secuencia de ADN por la poliadenilación y por una repetición de secuencia. Este c-ARN de larga longitud es el que servirá de plantilla para el genoma. El ARNm de larga longitud codifica la polimerasa y las proteínas de core HBcAg y HBeAg. Estas últimas son muy similares puesto que se traducen en el mismo marco de lectura desde dos codones de inicio diferentes. Dos ARNm más pequeños (2.4 y 2.1 bases) cuando se solapan codifican para las glicoproteínas de superficie. También hay un ARNm pequeño de 700 bases que codifica para una proteína que es una proteína cinasa y es un transactivador de la transcripción.
En el citoplasma, el c-ARN de longitud completa (3,500 bases) de sentido positivo es encapsidado por proteínas del núcleo (core). Dentro del core, el ARN se transcribe a ADN de cadena negativa vía la misma ADN polimerasa (transcriptasa inversa) que completó al ADN de cadena doble, y al mismo tiempo el ARN es degradado por una ribonucleasa H que es parte de la transcriptasa inversa. A diferencia de la transcriptasa inversa de los retrovirus, la transcriptasa inversa del VHB no requiere de un iniciador tARN. Más bien, la polimerasa misma actúa cono un iniciador y permanece unida covalentemente al terminal 5’ de la hebra de ADN de sentido negativo. Una proteína chaperona de la célula huésped, proteína de choque término 90, también es necesaria. La chaperona se asocia con la transcriptasa inversa permitiendo que se doble a una conformación activa.
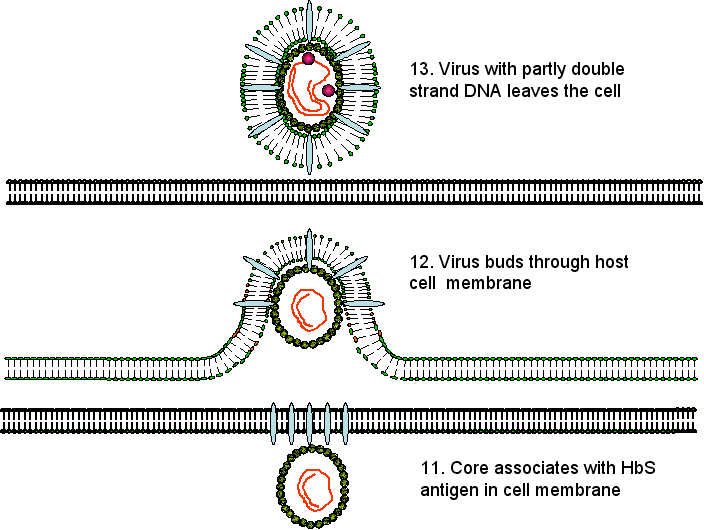
EL virus ahora yema a través del retícula endoplásmico y/o las membranas del cuerpo de Golgi (o quizás de un nuevo pre-Golgi compartimiento) de la célula huésped de donde adquiere el HBsAg. En esta etapa o después, la cadena negativa del ADN es parcialmente trascrita a una cadena positiva. Cuando la ADN polimerasa viral se usa para transcribir ARN a ADN, está actuando como una transcriptasa inversa parecida a la de los retrovirus; de hecho, la ADN polimerasa del VHB y la transcriptasa inversa de los retrovirus son muy similares, y puede que hayan evolucionado de un ancestro en común.
Las partículas víricas que contienen ARN o ADN en varios estadios de replicación pueden ser halladas en el torrente sanguíneo sugiriendo que la replicación de ácidos nucleicos no está muy controlada por el paso fuera de la célula. Además, las envolturas vacías embebidas con proteínas de membrana están siendo continuamente descamadas.
El problema
de la ARN polimerasa
Hay un claro inconveniente surgido del uso de la ARN polimerasa II del
huésped para transcribir el genoma de ADN viral a ARN (Refiérase a la
sección sobre los
retrovirus).
La función normal de la ARN polimerasa II es la de transcribir un gen a un
ARNm para su subsiguiente traducción a proteína. En el ARNm, todo lo que se
requiere es la información para sintetizar la proteína. En el gen de ADN,
hay información adicional presente que es necesaria para sintetizar ARN.
Esta información extra (que no es trascrita a ARN) incluye a la región
promotora (el sitio al que se une la ARN polimerasa), las regiones
potenciadoras que están corriente – arriba y corriente – abajo de la región
trascrita a ARNm y la región de poliadenilación. Por tanto un ARN mensajero
es más pequeño que el gen de ADN, aún cuando no hay intrones.
Los retrovirus superan la pérdida de información promotora/potenciadora como
resultado de usar la transcripción de la ARN polimerasa II llevando copias
internas de las regiones promotora y potenciadora (estas son las secuencias
U3 y U5 respectivamente). Estos virus duplican su promotor U3 interno y lo
transponen al terminal opuesto cuando el AND es trascrito a ARN. De igual
manera, los potenciadotes y otra información del terminal 3’ se almacenan
parcialmente en el interior (como regiones U5) y son transpuestos al
terminal opuesto. Estos sucesos dan lugar a las regiones de repeticiones
terminales largas (LTRs, por sus siglas en inglés) que sólo se encuentran en
la forma ADN del virus. Cuando la ARN polimerasa reconoce al promotor en la
región U3, encuentra el sitio de iniciación de la transcripción entre U3 y R
e inicia la transcripción en el comienzo de la región R. Esto conlleva a un
copiado leal del ARN original puesto que se pierden los terminales U3 y U5
(figura 4B).
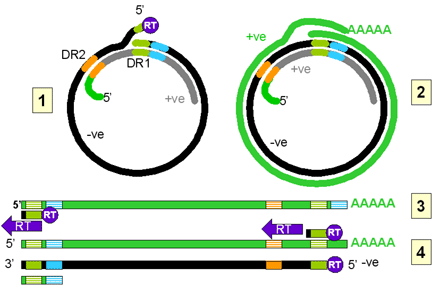
El mismo problema ocurre con los hepadnavirus que también tienen una forma
ADN de su genoma que es copiado a ARN por la ARN polimerasa II de la célula
huésped antes de copiar el ARN a ADN usando la transcriptasa inversa. No
obstante, el mecanismo es diferente, en este caso, la forma ADN del virus es
más pequeña que la forma ARN, justo lo opuesto de lo que ocurre en los
retrovirus.
Los hepadnavirus son virus pequeños de ADN y, en contraste a los retrovirus,
es el ADN que es empacado en la partícula vírica. Este ADN es copiado a ARN
en la célula infectada vía una ARN polimerasa II y el ARN resultando es
copiado de vuelta a ADN mediante una transcriptasa inversa en la partícula
vírica adulta.
En la partícula viral, el ADN sólo es parcialmente de doble cadena. La hebra
negativa está completa, pero no está ligada en un círculo. Hay terminales 5’
libres (con una molécula de transcriptasa inversa anexada) u terminales 3’.
El ADN se encuentra en forma de un círculo relajado puesto que está
hibridado a una copia parcial de la hebra positive. El ADN contiene dos
repeticiones directas (DR1 y DR2). DR1 está cerca del terminal 5’ de la
hebra negativa y DR2 está cerca del terminal 5’ de la semi-hebra positiva.
Al entrar al núcleo, la hebra negativa es ligada para formar un círculo
cerrado covalentemente. Este es posteriormente copiado por la ARN polimerasa
II del huésped. La polimerasa inicia más o menos 6 bases a la izquierda (en
la figura 4Ci-2) de la región DR1 y procede (en sentido de las manecillas
del reloj en la figura 4Ci-2) alrededor del círculo pasando las regiones de
iniciación y la DR1 y se detiene en la región de terminación/poli A (azul
pálido) que se encuentra más corriente abajo. El ARN se vuelve poliadenilado.
La copia de ARN es entonces más larga que el ADN circula covalentemente
cerrado (compare con lo que ocurre en los retrovirus) porque la región DR1
ha sido duplicada y una región poliA ha sido añadida.
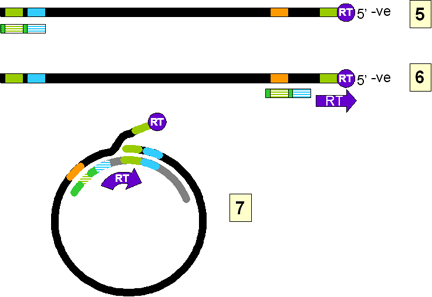
Este ARN se mueve al citoplasma en donde ocurre la encapsidación por
proteínas virales. Hay una señal para la encapsidación en la terminal 5’ del
ARN y por eso solo una molécula de ARN se encuentra en cada virión (compare
con la situación en los retrovirus). Ahora, en la partícula vírica misma, el
ARN es copiado a ADN usando la transcriptasa inversa. Todas las ADN
polimerasas necesitan un iniciador y en el caso de los retrovirus este es un
tARN de la célula huésped que es empacado en el virión. En los hepadnavirus,
la polimerasa es empacada en el virión, al igual que en los retrovirus,
aunque hay menor cantidad de polimerasas por virus en los hepadnavirus. La
misma transcriptasa inversa sirve de iniciador para la síntesis de hebras de
ADN de sentido negativo y permanece unido al terminal 5’ del ADN mediante un
residuo tirosina.
El ADN comienza en un grupo hidroxi de la tirosina usando, como plantilla,
una región adyacente al terminal 5’ del ARN (Fig. 4Ci-3). La polimerasa
copia por el DR1 cerca del terminal 5’ del ARN y termina al final de la
molécula de ARN. Luego, ocurre un intercambio de plantillas en el que el la
hebra negativa de ADN naciente se mueve a la región DR1 cerca del terminal
3’ (Fig. 4Ci-4). El por qué esto es necesario todavía no ha sido elucidado
porque la iniciación pudo haberse dado cerca de la región DR1 del terminal
3’. Desde DR1 del terminal 3’, el ADN se extiende acompañado por una
digestión de la hebra ARN que ha servido de plantilla llevada a cabo por la
RNasa H. La síntesis se detiene cuando se alcanza el terminal 5’ del ARN
(figura 4Ci-4). La hebra negativa ahora es redundante en su terminación. El
ARN no es completamente destruido y los últimos 15 nucleótidos permanecen
(figura 4Cii-5) para funcionar como iniciadores de la segunda síntesis de la
hebra de ADN (de sentido positivo). Este es transpuesto a la región DR2 del
terminal 5’ de la primera hebra de AND. Aquí ocurre entonces un cambio de
plantillas en el que la región DR1 del terminal 5’ de la hebra negativa es
remplazada por la región DR1 del terminal 3’, encirculando la plantilla
(figura 4Cii-7). La transcriptasa inversa luego copia alrededor del círculo
por una distancia variable para formar el ADN que se encuentra luego en las
partículas víricas maduras.
Carcinogénesis
Está claro que los individuos que son HBsAg positivos tienen un mayor riesgo de padecer de carcinoma hepatocelular que aquellos que son negativos para este antígeno. En los pacientes con hepatitis crónica, hay destrucción de hepatocitos como resultado de la respuesta del sistema inmune al virus. Esto provoca la regeneración (mediante división celular) de las células hepáticas lo que podría conllevar al cáncer. Aunque el virus no se integra durante el curso normal de la replicación, partes del genoma del VHB se encuentran integrados en el ADN de los pacientes con carcinoma hepatocelular. Esto puede resultar en la activación de un proto-oncogen celular de la misma manera que ocurre con muchos de los cánceres causados por retrovirus; de hecho, en la mayoría de los casos de carcinoma hepatocelular de las marmotas (un sistema modelo ampliamente utilizado), el ADN viral se encuentra cerca del myc o de un proto-oncogen similar. El carcinoma hepatocelular tarda años en manifestarse y esto puede ser reflejo de la infrecuencia de la integración en ausencia de la enzima integrasa. El tumor que se desarrolla es por tanto probable que sea un clon de una célula única en donde se ha dado este proceso.
Una proteína del VHB llamada proteína X es conocida por activar la src cinasa y esto también puede ser una causa de la carcinogénesis del VHB. Esta proteína puede también interactuar con el p53, uno de los genes supresores de tumores de la célula.
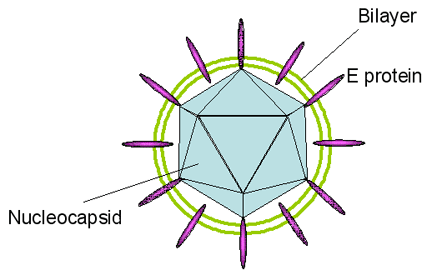 Figura 5
Figura 5Estructura del virus de la Hepatitis C
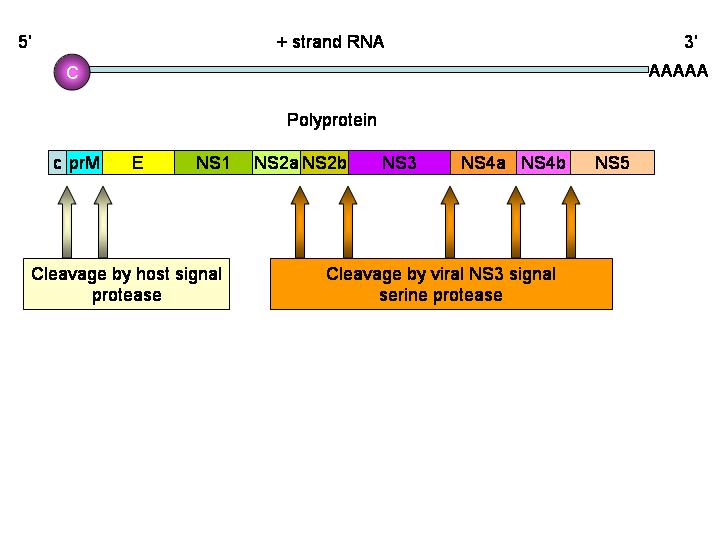 Figura 6
Figura 6
Procesamiento de la poliproteína del Flavivirus
VIRUS DE LA HEPATITIS C
El virus de la Hepatitis C es un flavivirus (de los cuales la malaria o fiebre amarilla es el prototipo) que causa hepatitis no-A no-B. Los Flavivirus (figura 5) son virus de ARN, icosaédricos, de sentido positive y obtienen su envoltura de la célula huésped. La partícula vírica mide cerca de 30 a 60nm de largo. El genoma de 9,100 bases codifica para diez proteínas. En muchas maneras, los flavivirus son parecidos a los picornavirus con la prominente excepción de que son envueltos. El ARN viral tiene una chapa 5’ pero no tiene un tracto 3’ poli A. Al igual que con los picornavirus, hay un producto proteico de un marco de lectura abierto. La poliproteína es clivada por una actividad proteasa viralmente codificada. A diferencia de los picornavirus, la proteína naciente contiene una secuencia de señal que resulta en la traducción de la unión de ribosomas a la superficie citoplasmática del retículo endoplásmico. La proteína de envoltura (E) por tanto atraviesa y se incrusta en la membrana y se secuencia de señal es removida por un proteasa celular. El resto de la proteína es removida de la proteína E y por ellos se vuelve citoplásmica. Esta proteína es dividida en proteínas NS1, NS2, NS3 y NS4. Las NS2 y NS4 son cortadas de nuevo (para dar NS2a, NS2b, NS4a y NS4b) (figura 6). El VHC se une tanto al antígeno CD81 como a los receptores de lipoproteína de baja densidad (LDL) en los hepatocitos vía su glicoproteína E2. También hay evidencia de que puede unirse a glucosaminoglicanos.
 Figura 7
Figura 7Agente de la Hepatitis Delta CDC
ANTÍGENO DELTA DE LA HEPATITIS
La Hepatitis D (figura 7) es un virus altamente defectuoso puesto que no puede producir visiones infecciosos sin la ayuda de un virus ayudante co – infectante. Este virus ayudante es el virus de la hepatitis B que suple el antígeno HBsAg de superficie. Cuando yema fuera de la célula, el VHD adquiere una membrana que contiene HBsAg. EL VHD es similar a un viroides vegetal puesto que es un genoma de ARN circular pequeño (1,700 bases) pero a diferencia de los viroides vegetales, el ARN codifica una proteína llamada antígeno delta. Esto forma un complejo con el ARN. El ARN es de cadena sencilla y sentido negativo y es un círculo covalentemente cerrado. Dada la gran cantidad de apareamiento de bases, el ARN se vuelve una estructura baciliforme (figure 8).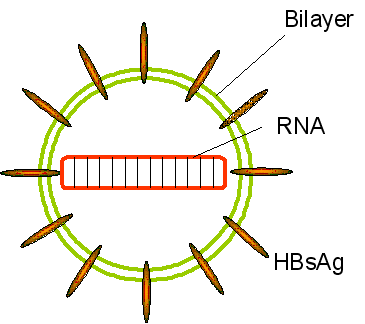 Figura 8
Figura 8Agente Delta de la Hepatitis - estructura
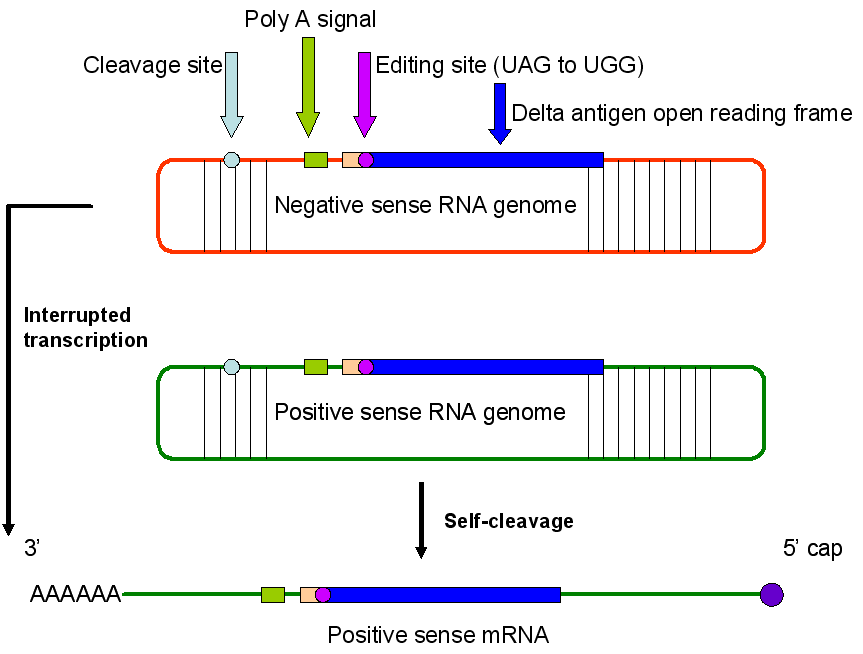 Figura 9
Figura 9
Agente delta de la Hepatitis. Tres formas de ARN.
Adaptado de Wagner y Hewitt.: Virología Básica. Publicaciones Blackwell
El VHD solo puede formar una partícula infecciosa si la célula en la que se replica está co-infectada con el VHB puesto que este último provee el antígeno de superficie HBsAg que es necesario para reinfectar otra célula. El HBsAg del VHD se una al mismo receptor de superficie que el VHB y el virus se funde con la membrana celular. El tropismo del VHD es por tanto el mismo del del VHB. El genoma de ARN está cubierto con el antígeno delta, la única proteína codificada por el ARN. El antígeno delta, el cual es expuesto cuando se pierde la envoltura, tiene una señal de localización nuclear que señala al genoma del núcleo. Aquí el genoma es copiado por la ARN polimerasa II de la célula huésped, la enzima normalmente sintetiza ARNm. La ARN polimerasa II es utilizada por algunos otros virus para copiar sus genomas, por ejemplo, los retrovirus, pero en ese caso la polimerasa copia ADN a ARN (lo cual es la función normal de la enzima en una célula no infectada). En la replicación del VHD, la polimerasa copia ARN a ARN. El sentido negativo del genoma del ARN es copiado a una hebra positiva de forma tal que también es circular. El ARN genómico también puede ser trascrito a un ARNm lineal chapado en su terminal 5’ y poliadenilado en su terminal 3’, el cual es más pequeño que el ARN genómico y contiene los pequeños marcos de lectura abiertos desde los que es traducido el antígeno delta; o este puede ser generado de ARN circular positivo por un proceso auto catalítico que cliva el ARN. Por ende, el ARN actúa como una ribozima, que es un catalítico de ARN (figura 9).
El antígeno Delta, traducido de un ARNm tiene dos formas que difieren en tamaño por 19 aminoácidos (195 residuos vs. 214 residuos). La formación del antígeno delta mayor sucede por un extraño mecanismo en el cual una enzima de la célula huésped llamada adenosina deaminasa activada por ARN de cadena doble convierte el codón UAG (de finalización) e un codón UGG que permite que el proceso de traducción se prolongue hasta el próximo codón de finalización. El antígeno delta menor está implicado en la replicación del genoma pero el mayor suprime la replicación. Esto conlleva a la promoción del ensamblaje de partículas víricas.
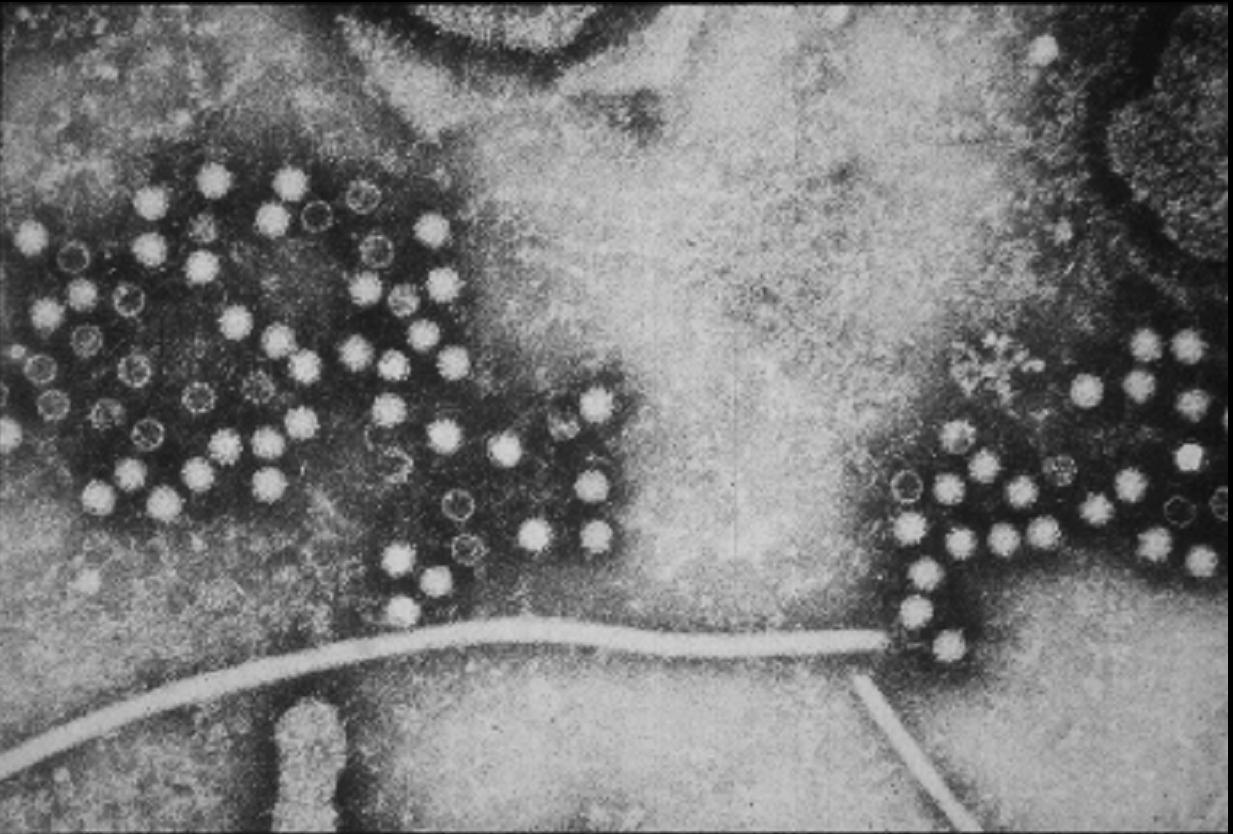 Figura 10
Figura 10Virus de Hepatitis E CDC
VIRUS DE LA HEPATITIS E
Este virus (figura 10), el cual causa hepatitis entérica no A no B, parece estar relacionado con los Calicivirus pero su clasificación aún no está definida puesto que la organización de su genoma no es como la de los Caliciviridae. En su secuencia, el VHE es más parecido a la rubella que es un Togavirus. El VHE es un pequeño (aproximadamente 34nm) virus de ARN, redondeada, icosaédrico, de sentido positivo que no posee envoltura. Tienen una superficie más bien lisa pero no tanto como la del VHA. El genoma tiene un tracto de poliA y está chapado en el terminal 5’. Hay tres marcos de lectura abiertos que se solapan; cada uno es en un marco codificador diferente. Basándose en motivos de secuencia, el marco de lectura abierto 1 (ORF1, por sus siglas en inglés) aparente tener varias actividades enzimáticas. Estas pueden involucrarse en el chapamiento del ARN, la proteolisis y en actividad de ADN polimerasa ARN dependiente. El ORF2 es la proteína estructural y puede estar glicosilada. Parece tener una secuencia de señal sugestiva de que su proteína codificada puede entrar al retículo endoplásmico. El tercer ORF codifica una fosfoproteína de función desconocida que interactúa con el citoesqueleto de la célula huésped. No se conoce mucho de la replicación del VHE pero es probable que la hebra positive de ARN sea copiada en un intermediario de hebra negativa por una polimerasa viral.
VIRUS DE LA HEPATITIS G
El virus de la Hepatitis G es un flavivirus, al igual que el VHC con el que está cercanamente relacionado. Se asocia con algunos casos de hepatitis no-A, no-B, no-C, no-D agudos o crónicos. Aunque parece ser común en la sangre humana, puede no se una causa significativa de hepatitis en los humanos.|
Tabla 1 |
|||||
| Hepatitis A | Hepatitis B | Hepatitis C | Hepatitis Delta | Hepatitis E | |
| Familia viral | Picornavirus | Hepadnavirus | Flavivirus | ARN Circular similar a viroides vegetales | Similar a Calicivirus |
| Ácido nucleico | ARN (sentido +) | ADN (parcialmente de cadena doble) | ARN (sentido +) | ARN (sentido-) | ARN (sentido+) |
| Patología causada | Hepatitis infecciosa | Hepatitis sérica | Hepatitis No-A, no-B | Hepatitis entérica no-A, no-B | |
| Tamaño | ~ 28nm | ~40nm | 30 - 60nm | ~ 40nm | 30 - 35 nm |
| Envoltura | No | Sí | Sí | Sí | No |
Regreso a la sección de Virologia Microbiología e Inmunología on line
Derechos de autor 2008 The Board of Trustees of the University of South Carolina
Esta página se modificó recientemente en Monday, March 03, 2008
Mantenimiento de la pagina por Richard Hunt
Favor de reportar problemas a rhunt@med.sc.edu